
STRUCTURA SECUNDARA SI TERTIARA A ARN
Moleculele de ARN, care contin doua catene imperechiate, reprezinta o exceptie care este intalnita la cateva virusuri. Diferitele tipuri de ARN sunt, de fapt, produsi ai reactiilor de transcriptie a unei singure catene. In acest caz structura primara, reprezentata de compozitia si secventa nucleotidica constituie:
a) suportul copiei codificate de gene destinat traducerii in proteine;
b) obiectul unei serii de modificari importante si variate care determina obtinerea unor baze atipice (dihidro-uridina, pseudo-uracilul, etc.);
c) structura care individualizeaza situsuri specifice de legare a proteinelor;
d) structura care determina orientarea spatiala a polimerului la nivel secundar si tertiar.
Structura secundara a ARN
Fortele de stivuire si de imperechere a bazelor determina si in cazul ARN, structuri sub forma de elice. Acestea nu au perfectiunea dublului helix de referinta prezent in structura ADN, dar impreuna cu regiunile existente intre ele se organizeaza in structuri definite prin notiunea de motive.
Elicele ARN
Numai sub actiunea fortelor de stivuire, scheletul monocatenei tinde sa ia o forma de elice dreapta simpla si neregulata. Tendinta de apropiere dintre doua purine este mai puternica, astfel incat daca doua purine sunt separate de o pirimidina ele pot determina indepartarea acesteia pentru a se stivui. Cu toate acestea, conformatia dominanta este cea de dubla elice care se poate obtine din: i) doua catene de ARN de tipul celor descrise mai sus; ii) dintr-o catena ADN si una ARN in cazul hibrizilor tranzitorii care apar in celula in timpul transcriptiei; iii) doua segmente de ARN distante care prezinta o complementaritate destul de importanta pentru a se imperechea.
Daca comparam aceasta structura cu dublul helix ADN constatam urmatoarele: a) catenele sau segmentele sunt de asemenea antiparale (antisens);
b) imperecherile standard (A-U si G-C) sunt imbogatite in cazul ARN cu alte imperecheri Watson-Crick (cuplul G-U) dar si de imperecheri ne-standard de tip A-I, U-I si C-I foarte frecvente in cazul moleculelor ARNt. De retinut ca afinitatea relativa a perechilor G-C, A-U si G-U este 1 000, 100, 1(Figura 4.1
Figura
4. 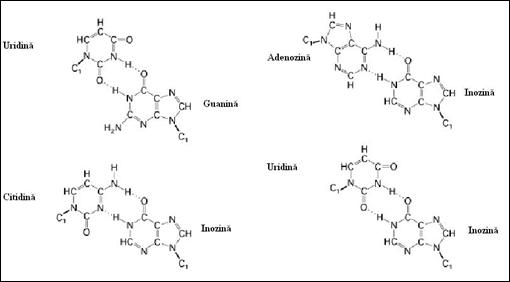 Tipuri de legaturi de hidrogen care sunt specifice
structurii secundare a ARN (dupa Molecular Cell Biology Lodish H.,
et. al., 2000)
Tipuri de legaturi de hidrogen care sunt specifice
structurii secundare a ARN (dupa Molecular Cell Biology Lodish H.,
et. al., 2000)
c) apar baze modificate derivate atat de la bazele purinice cat si pirimidinice (Figura 4. 2
d) radicalii hidroxil din pozitia 2' a ribozei se opun, prin interferenta lor puternica cu baza, rasucirii pentru a forma o conformatie B. Inclinarea pe care acestea o impun planului bazei (de aproximativ 200 fata de axa) are drept rezultat aparitia unei conformatii asemanatoare cu conformatia A (numita adesea ARN 11 pentru a accentua faptul ca sunt necesare 11 pb/tur).
Intr-o molecula de ARN monocatenara, imperecherea bazelor se face pe lungimi foarte variabile: de la una la mai multe sau la mai multe zeci de perechi de baze. Diferitele elice, formate impreuna cu segmentele aleatoare ne-imperecheate care le separa, desemneaza cateva structuri bine definite si reproductibile care reprezinta gradul secundar de organizare. Aceste motive pot fi reprezentate printr-un grafic bidimensional.
Motivele elementare 2-D
In figura 4.3 a este prezentata structura secundara a uneia dintre cele mai cunoscute molecule de ARN. Sunt evidentiate motivele identice care se regasesc in structura tuturor moleculelor de ARN de transport. Se observa prezenta unor "proeminente" si "pliuri" care determina orientarea segmentelor. Structura reprezinta o alternanta interesanta de tije, la nivelul carora bazele se imperecheaza pe baza de complementaritate, si bucle.
Buclele difera prin: i) pozitia lor interna sau externa in organizarea generala a moleculei; ii) marimea elementelor lor, tija si bucla; aceasta variaza de la mai multe sute pentru cele mai mari la cateva unitati pentru buclele cu motive in agrafa, mai putin perfecte decat cele descrise in cazul ADN.
Aceste motive sub forma de agrafa sunt importante in structura si functionarea ARN. Buclele acestora sunt constituite din secvente particulare: i) secvente stabilizante cum sunt UNCG care determina formarea unor bucle solitare; ii) secvente dotate cu potential de interactiune cu alte secvente, cum este cazul buclelor GNRA si a buclelor de sapte baze care sunt caracteristice ARNt.
Structura tertiara a ARN
Interactii intre structurile secundare
Diferite regiuni secundare, mai mult sau mai putin distantate in reprezentarea plana, determina prin interactie aparitia de replieri spatiale. Legaturile implicate sunt legaturi de hidrogen:
a) formate intre bazele din regiunile ne-imperecheate care determina perechi Watson-Crick sau alte tipuri;
b) determinate prin participarea bazelor modificate (Figura 4.2
c) foarte originale care implica grupari ale scheletului care formeaza asocieri cu trei sau chiar patru parteneri. In aceste cazuri pot participa gruparile hidroxil din pozitia 2' a ribozei sau un atom de oxigen al gruparii fosfat.
Caracteristicile structurilor 3-D
In urma interactiunilor descrise mai sus iau nastere structurile 3-D. Un numar restrans de asocieri simple, prezente in motivele arhitecturale 3-D, au fost deja identificate iar altele sunt in curs.

Figura 4. Tipuri de baze modificate prezente in structura acizilor nucleici
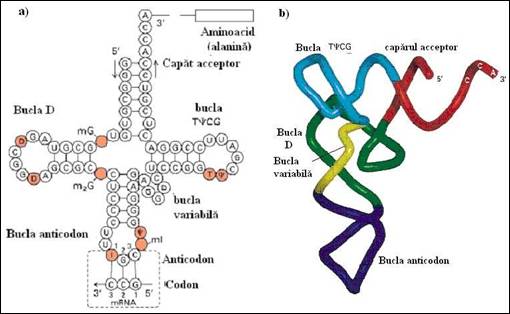
Figura 4. Structura ARN de transport: a) Motivele elementare de structura 2-D; b) Structura tertiara in forma de L (dupa Molecular Cell Biology Lodish H., et. al., 2000).
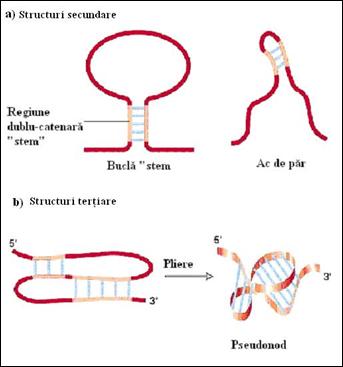
Figura 4. Tipuri de structuri secundare si tertiare caracteristice moleculelor de ARN: a) structuri secundare; b) structuri tertiare si pseudo-tertiare (dupa Molecular Cell Biology Lodish H., et. al., 2000)
Pana in prezent au fost caracterizate cateva motive arhitecturale:
a) doua elice care se alatura sau se suprapun partial (imbuca) prin intermediul fosei lor aplatizate;
b) o elice fixata intr-o fosa: i) un segment ne-imperecheat se fixeaza prin imperecheri de tip Hoogsteen la nivelul fosei profunde si prin intermediul ribozei la nivelul altui fragment: se formeaza astfel o tripla elice; ii) sau prin prinderea unei bucle GNRA in fosa sa aplatizata;
c) asocieri intre regiuni ne-imperecheate ale unei bucle cu un segment aleator determinand producerea unor pseudo-noduri (noduri false) (Figura 4.4
Organizarea in domenii
Conformatia globala a moleculelor de ARN mari prezinta domenii independente care determina aparitia unei structuri specifice si compacte pentru unele din aceste motive. Acestea sunt separate de fragmente flexibile. Cel mai bun exemplu, in acest sens, il constituie structura secundara a ARNr 16 S, de 1542 nucleotide, de la E. coli (Figura 4.5). Aceasta a fost obtinuta prin compararea secventelor de la mai multe tulpini si se considera a fi conservata evolutiv. In figura 4.5 sunt prezentate cele patru domenii si localizarea principalelor subdomenii la nivelul subunitatii ribozomale 30 S. Fiecare domeniu este alcatuit din structuri de tip tija bucla specifice.
Motive secundare si tertiare, domenii.. Poate fi asemanata organizarea structurala a ARN cu a proteinelor?
Asemeni proteinelor, secventa nucleotidica a polimerului contine intreaga informatie a conformatiei sale. In acelasi mod, achizitionarea structurii spatiale este o tema inca actuala de cercetare, care vede confruntandu-se doua conceptii opuse: fie motivele se formeaza secvential si paralel cu biosinteza catenei fie, din contra, molecula completa suporta o pliere aleatoare provizorie la nivelul careia se realizeaza ajustarea motivelor.
Indiferent de cronologie, important este ca o molecula de ARN poate exista sub diferite forme conformationale reversibile, dependente de ioni magneziu sau de liganzi de tipul polipeptidelor sau proteinelor.
In cazul ARN, cristalografia cu raze X este adesea un esec deoarece densitatea lor de sarcina reprezinta un impediment pentru determinarile cristalografice sau determina rupturi. Utilizarea tehnicii RNM este limitata la moleculele de marime mica. Pentru moleculele ARN de marime mare, metoda cea mai buna pentru identificarea replierilor 2-D si 3-D, plecand de la secventa primara, o constituie modelarea moleculara.
Mult timp, modelele de studiu ale conformatiei 3-D au fost constituite din moleculele de ARN de transfer (ARNt). In prezent, ribozimele de tipul celei descoperita la Tetrahymena reprezinta un material de studiu foarte interesant.
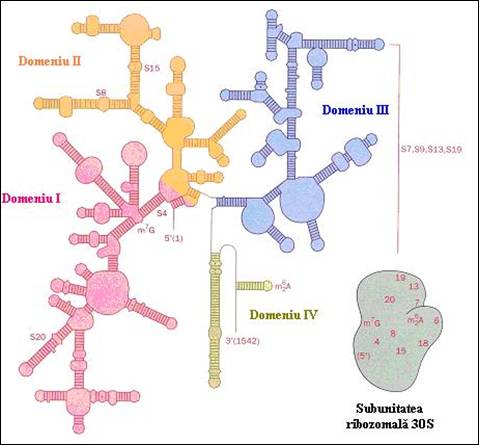
Figura 4. Structura secundara a ARNr de 16 S de la E. Coli de 1542 nucleotide bazata pe compararea secventelor de la diferite tulpini (dupa Biochemistry, Voet&Voet, 1995)
Tipuri de ARN
Studiul aprofundat al structurii diferitelor molecule de ARN nu prezinta interes real decat in corelatie cu functiile acestora in mecanismul transcriptiei sau in cadrul biosintezei proteice. In aceste conditii ne vom limita numai la cateva aspecte ilustrative privind structurile elementare descrise anterior.
ARN mesager
Reprezinta copii ale genelor ADN. Aceste amprente in limbaj genetic ternar (aminoacizii sunt codificati de triplete de baze numite codoni) contin secventa proteinei care va trebui sintetizata, flancata de secvente aditionale necesare functionarii si reglarii masinariei de translatie. Conceptul de mesager a fost formulat pentru prima data de F. Jacob si J. Monod in 1961. Acestia au primit impreuna cu A. Lwoff premiul Nobel pentru fiziologie si medicina in 1965.
La procariote informatia este adesea copiata de pe operonii bacterieni, ARNm fiind o molecula policistronica (Figura 4.6a
La eucariote ARNm este monocistronic si contine exoni si introni. Utilizarea ARN mesager permite celulei sa separe stocarea si utilizarea informatiei. In nucleul eucariotelor, in urma procesului de transcriptie, moleculele de ARN nuclear heterogen (ARNnh) formeaza o colectie de transcripti pentru numeroase gene nucleare: i) transcripti primari, marimea lor este identica cu cea a genei; ii) molecule partial maturate care sunt lipsite de un numar variabil de introni. Moleculele de ARNm citoplasmatic sunt produsul unei astfel de colectii complexe de precursori care au suferit un complex proces de maturare (Figura 4.6b). Precursorii ARNm de la eucariote sunt procesati inainte de transportul ARNm in citoplasma unde urmeaza a fi tradus de catre ribozomi. Acest proces consta in: i) adaugarea structurii "cap"; ii) scindare si poliadenilare in pozitia 3'; iii) inlaturarea intronilor si sudarea exonilor ("splicing").
In timp ce genele raman izolate in nucleu, unde sunt parti integrante ale moleculelor enorme de ADN, informatia acestora poate fi comunicata unei molecule de ARN mobila, mult mai mica si capabila sa treaca in citoplasma. La acest nivel, molecula serveste drept model pentru a dirija incorporarea aminoacizilor intr-o ordine specifica, codificata de secventa nucleotidelor in ADN si ARNm. De asemenea, folosirea ARNm permite celulei sa isi amplifice puternic activitatea. O molecula de ADN poate servi de model pentru producerea unui numar mare de molecule de ARN care, toate vor reprezenta matrite pentru producerea unui numar si mai mare de catene polipeptidice.
Durata de viata a moleculelor de ARN mesager este scurta. Din acest punct de vedere, ARNm este opusul ARNt a carui durata de viata este mult mai mare. Aceeasi molecula de ARNt este folosita de mai multe ori pentru a asigura transportul aminoacidului corespunzator la nivelul ribozomilor. La bacterii durata de viata a unui ARNm este de numai cateva minute. La eucariote aceste molecule sunt mai stabile durata de viata variind de la cateva minute la cateva zile.
ARNm se reinnoieste foarte repede, aceste molecule fiind permanent produse si degradate. Ele au, asemeni trandafirilor, viata unui mesaj. Cu toate acestea, aceeasi molecula de ARNm poate fi citita de mai multe ori la nivelul ribozomilor.
Din punct de vedere cantitativ ARNm nu reprezinta decat cateva procente din totalul moleculelor de ARN din celula.
In timpul transcriptiei unui fragment de ADN, in molecula de ARN corespunzatoare, se formeaza tranzitoriu hibrizi scurti ARN-ADN cu conformatie A, la nivelul buclei unde catenele de ADN sunt local desfacute.

Figura 4. Sinteza moleculelor de ARNm: a) transcriptia ARNm policistronic la procariote; A, B; C, D si E sunt genele operonului triptofan (trp)
b) transcriptia si maturarea ARNm la eucariote (gena b globinei); ARN transcris de la nivelul unei unitati complexe (albastru) contine exoni (c) si introni (d); poate fi procesat pe mai multe cai pentru a conduce la unul sau la mai multe unitati ARNm monocistronice functionale; liniile punctate marcheaza inlaturarea intronilor (dupa Molecular Cell Biology Lodish H., et. al., 2000)
Etapa de descifrare, pas cu pas, a mesajului purtat de ARNm de catre ribozomi, pentru traducerea acestuia in catena polipeptidica, este reglata de motive in forma de agrafa (ac de par) sau, in anumite cazuri, de pseudo-noduri tertiare (Figura 4.4). Informatia genetica copiata in structura ARNm, in urma procesului de transcriptie, este tradusa in structura proteinelor prin intermediul codului genetic care realizeaza traducerea mesajului dintr-un cod format din inlantuirea repetitiva si aleatoare a patru nucleotide in altul, reprezentat de alternanta a 20 de aminoacizi. Fiecare aminoacid este codificat de una sau mai multe secvente de trei baze, numite codoni, din structura ARNm. Codul genetic aste format din 64 de combinatii posibile (codoni) dintre care 61 specifica amino acizi si trei (AAA, AAG si AGA) semnalizeaza incheierea traducerii (STOP). Codonul de initiere, AUG codifica formil-metionina la procariote si metionina la eucariote. Codul genetic este degenerat deoarece majoritatea aminoacizilor sunt codificati de mai multi codoni (Figura 4. 8). Codul genetic standard este comun pentru procariote si eucariote. Mici diferente au fost evidentiate in cazul codului genetic utilizat de catre mitocondrii.
ARN de transfer
Aceste molecule au rolul de a adapta fiecarui codon, prezent in structura ARNm, aminoacidul corespunzator. Sunt molecule polinucleotidice mici de 75-90 de resturi a caror functionalitate se bazeaza pe existenta a doua situsuri strict specifice: i) capatul 3' care asigura fixarea unuia din cei 20 de aminoacizi, intr-o forma activata; ii) capatul anticodon, structura complementara codonului care se potriveste foarte bine pe structura ARNm prin imperechere complementara. O molecula de ARN de transfer, incarcata cu un aminoacid, poate fi reprezentata conform schemei elementare din figura 4.7.
O molecula de ARNt are o serie de caracteristici datorate structurii sale primare:
a) contine nucleotide atipice, neobisnuite, prin natura bazelor pe care le contin. Astfel, in structura ARNt gasim hipoxantina al carei nucleotid corespunzator este IMP (Figura 4.9
b) prezenta timinei, o baza specifica ADN si a altor baze metilate.
Deoarece aceste baze nu sunt incorporate in momentul sintezei ARN, ele sunt obtinute prin modificarea secundara a uneia dintre cele patru baze, A, U, C, G, intalnite in mod normal in structura primara a ARN. Astfel, IMP provine din deaminarea adeninei din AMP si TMP provine prin metilarea UMP. Aceste modificari se numesc post-transcriptionale.

Figura 4. Schema unei molecule de ARNt incarcata: a) schema generala a structurii secundare a moleculei; b) modul de formare a legaturii intre ARNt si aminoacil (dupa Biochemistry, Voe t& Voet, 1995)

Figura 4.8. Codul genetic standard. Codonul AUG care codifica pentru metionina este cel mai comun codon start. Trei dintre codoni AAA, AAG si AGA functioneaza drept codoni STOP. Se poate observa degenerescenta codului genetic deoarece numai metionina si triptofanul sunt codificati de un singur codon, restul aminoacizilor fiind codificati de doi la sase codoni.
Prima molecula de ARNt, secventializata in 1965, a fost cea pentru alanina de la drojdii (ARNtAla, 76 nucleotide). De atunci alte mii de molecule de la peste 500 de organisme sau din organite diferite au fost secventializate. Compozitia lor se caracterizeaza printr-o varietate si un procentaj mare de baze modificate (Figura 4.3). Analiza tridimensionala, prin difractie cu raze X, a ARNt pentru fenilalanina cristalizat, nu a fost realizata decat 10 ani mai tarziu. Din pacate, de atunci numai un numar destul de mic de molecule ARNt au fost analizate prin aceasta tehnica.
Rezultatele diferitelor metode de analiza sau a modelelor conformationale sunt uimitoare: in ciuda diferentelor existente intre secventele nucleotidice primare, din punct de vedere al structurii secundare si tertiare, moleculele de ARNt se prezinta sub forma aceluiasi model structural.
Structura secundara sub forma de "frunza de trefla"
Imaginea vine de la reprezentarea in plan a moleculei (Figura 4.7a): o tija cu trei foi de trefla, a patra putand fi adesea numai intrezarita.
Structura in care reperam motivele descrise mai sus, se compune din patru brate elicoidale pe care le diferentiem prin functia si caracteristicile lor structurale:
1) extremitatea 3' CCA (constituita din trei nucleotide CMP, CMP si AMP) reprezinta tija care constituie bratul acceptor al aminoacidului. Acesta se leaga covalent la hidroxilul extremitatii 3' hidroxil (Figura 4.7b
2) bratul purtator al anticodonului: este o structura sub forma de agrafa a carei bucla formata din sapte nucleotide poarta anticodonul. Numim anticodon grupul de trei nucleotide care este situat la nivelul unei bucle a ARNt si are capacitatea de a se imperechea prin legaturi slabe cu codonul intr-o maniera antiparalela si complementara. In cazul codonului si anticodonului este absolut necesara precizarea sensului de scriere a secventei. Astfel, anticodonul "3' UAC 5' " corespunde codonului "5' CAU 3'" (Figura 4.8
3) bratele D si TYC isi datoreaza denumirile si simbolurile faptului ca buclele lor contin baze modificate rare: pentru bratul D este dehidrouridina iar pentru TYC, este prezenta pseudouridina pozitionata intre timina si citozina si notata Y Figura 4.7a
Sistemul de numerotare de la 5' la 3' (de la stanga la dreapta) a fost utilizat pentru prima data in cazul structurii ARNt de 76 nucleotide si a fost adoptata prin conventie pentru reprezentarea normala a unei molecule ARNt.
Diferente intre structurile secundare ale diferitelor molecule pot sa apara la nivelul buclei bratului D dar acestea nu depasesc insa 4 baze. De asemenea, unele molecule poseda o bucla suplimentara, care reprezinta regiunea cea mai variabila a moleculelor de ARNt. In functie de structura, la acest nivel distingem: i) moleculele de ARNt de clasa 1, cele mai bine reprezentate si care prezinta un brat suplimentar scurt
de 3 la 5 nucleotide; ii) moleculele ARNt de clasa 2 care poseda un brat suplimentar lung de 13 la 21 nucleotide dintre care 5 sunt complementare (Figura 4.10
Imperecherea secundara a bazelor determina formarea de elice constante. Daca parcurgem structura din directia 5' catre 3' numarul de perechi de baze din care acestea sunt formate este 7, 3 (sau 4), 5 si 5. Imperecherile bazelor sunt de tip Watson-Crick cat si de alte tipuri.
Compararea secventelor moleculelor de ARNt evidentiaza conservarea in pozitii invariabile a unor baze pirimidinice sau purinice.
Structura tertiara in L a moleculelor ARNt
In figura 4.3b este prezentata structura tertiara a unei molecule de ARNt. Numarul mare de punti de hidrogen, care se formeaza intre baze situate la distanta, realizeaza replierea moleculei si aparitia unei conformatii cu doua regiuni elicoidale ale caror axe sunt situate in planuri perpendiculare.
Una dintre aceste elice lungi se formeaza intre bratul D si anticodon iar cealalta intre bratul acceptor si TYC. Cu toate ca molecula in aceasta conformatie 3-D este compactata, constatam ca doi poli sunt particular accesibili: extremitatea 3' si bucla anticodon.
In ciuda numarului mic de cristalizari reusite pentru moleculele de ARNt, datele obtinute sunt suficiente pentru a admite existenta unei structuri identice sau foarte apropiate pentru toate tipurile de ARNt.
ARN ribozomal
Ribozomi
Ribozomii reprezinta sediul sintezei proteice si sunt agregate multi-moleculare necovalente, mari si compacte de proteine si ARN. Ei sunt formati din doua subunitati diferite, ca marime si compozitie, care in cazul micsorarii concentratiei de ioni de Mg2+ se separa reversibil. La baza intelegerii structurii si functiei ribozomilor stau studiile biochimice realizate de Nomura cat si utilizarea tehnicilor imunologice si de microscopie electronica.
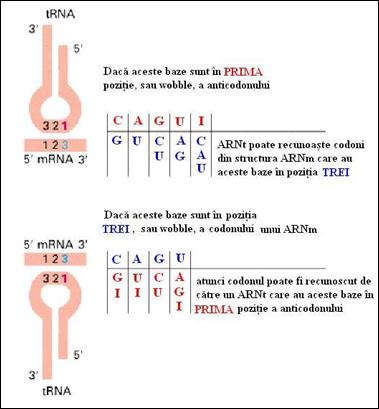
Figura 4. Recunoasterea codon anticodon (dupa Molecular Cell Biology Lodish H., et. al., 2000)
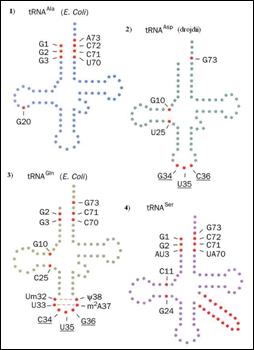
Figura 4.10. Tipuri de molecule de ARNt diferentiate in functie de marimea bratului suplimentar: structurile 1, 2 si 3 apartin categoriei clasei 1 in timp ce 4 apartine clasei 2. Sunt marcate elementele de identitate majore in cele patru tipuri. Fiecare baza din structura ARNt este reprezentata de cercuri pline. Cercurile rosii indica pozitiile identice pentru recunoasterea ARNt de catre aminoacil-ARNt sintetaza. Bazele din structura anticodonului care reprezinta elemente de identificare sunt subliniate.(dupa Biochemistry, Voet&Voet, 1995)
Tratarea subunitatilor cu agenti denaturanti (fenol, uree, sulfat de amoniu) disociaza legaturile slabe si precipita proteinele determinand separarea constituentilor ARN, in trei specii diferite, si proteici, in mai multe zeci de proteine diferite.
Particulele ribozomale din citoplasma eucariotelor superioare sunt mai mari decat cele prezente la procariote. Celula ai carei ribozomi au fost cel mai mult studiati este E. coli. Compozitia si caracteristicile acestor ribozomi sunt date in figura 4.11. Ribozomii 70S ai E. coli sunt formati din doua componente: o subunitate mare (50S) si o subunitate mica (30S). S este prescurtarea pentru unitatea Svedberg care reprezinta masura vitezei de sedimentare prin ultracentrifugare. Coeficientul de sedimentare depinde nu numai de masa dar si de rigiditatea particulei. Astfel, se explica de ce asamblarea subunitatilor 50S si 30S poate da particule caracterizate printr-o constanta de sedimentare de 70S. Fiecare dintre aceste subunitati sunt formate dintr-un amestec de proteine, denumite proteine ribozomale si ARN ribozomal (ARNr).
Subunitatea 30S contine: i) o molecula ARNr 16S de 1 542 nucleotide; ii) 21 de proteine diferite, prezente intr-un singur exemplar. Proteinele au fost denumite de la S1, S2 la .S21. In cazul proteinelor ribozomale S are semnificatia "small" si desemneaza proteinele care intra in constitutia subunitatii mici. Ansamblul ARNr impreuna cu proteinele ribozomale are o masa de aproximativ 900 kDa.
Subunitatea 50S contine: i) o molecula ARNr 5S de 120 nucleotide; ii) o molecula ARNr 23S de 2 904 nucleotide; iii) 34 proteine ribozomale denumite L1, L2, L31. L este folosit pentru "large" si semnifica prezenta acestor proteine in constitutia subunitatii mari. Treizeci dintre aceste proteine se gasesc intr-un singur exemplar iar una dintre proteine in patru exemplare. Ansamblul proteine-ARN al subunitatii mari are aproximativ 1,6 milioane daltoni.
In momentul asamblarii subunitatii mici cu cea mare se formeaza intre cele doua o fosa la nivelul careia va trece ARNm in timpul traducerii sale in proteine.
La eucariote ribozomii sunt mai mari, 80S, cele dupa subunitati fiind de 40S si respectiv 60S. Compozitia acestora este diferita in ceea ce priveste tipurile de ARNr si numarul proteinelor din care acestia sunt constituiti. Subunitatea mica contine o molecula de ARNr 18S si 33 de proteine ribozomale iar subunitatea mare trei molecule ARNr (5S, 5,8S si 28S) si 45 proteine ribozomale.
Aceste diferente ale constitutiei ribozomilor prezinta un interes foarte mare deoarece anumite antibiotice, care actioneaza la nivelul ribozomilor, pot sa aiba o actiune specifica asupra ribozomilor bacterieni fara a afecta ribozomii proprii organismului gazda (Figura 4.13

Figura 4. Principalele caracteristici ale ribozomilor de la procariote si eucariote (dupa Molecular Cell Biology Lodish H., et. al., 2000)
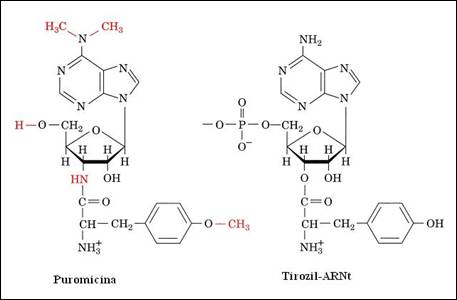
Figura 4. . Structura puromicinei analoga cu tirozil ARNt
Alte antibiotice de tipul puromicinei intervin in procesul de sinteza proteica atat la eucariote cat si la procariote determinand terminarea prematura a sintezei catenei proteice datorita analogiei structurale. Aceasta molecula este asemanatoare cu extremitatea 3' a unui ARNt incarcat (Figura 4.12) si poate ocupa situsul aminoacil de pe structura ribozomului. Pozitionarea sa permite transferul catenei in formare pe puromicina prin formarea unei legaturi covalente. Fixarea moleculei de antibiotic la capatul carboxi-terminal al polipeptidului stopeaza continuarea sintezei proteice.
Structura ARN ribozomal
Moleculele de ARNr sunt constituenti majori ai ribozomilor reprezentand aproximativ 65% din structura acestora.
Moleculele ARNr se caracterizeaza prin cateva aspecte structurale importante care le diferentiaza de celelalte tipuri de ARN. Acestea sunt molecule care sufera metilare dupa sinteza polinucleotidului la nivelul adeninei (N-dimetil adenina) sau prin formarea de O-metilriboze. Metilarea hidroxilului din pozitia 2' a ribozei protejeaza polimerul de hidroliza pontilor fosfodiester si prelungeste durata de viata a acestuia.
A fost determinata structura primara a moleculelor de ARNt de diferite origini. Analiza secventelor si studiile realizate asupra structurilor secundare an relevat acelasi paradox ca si in cazul ARNt, si anume evidentierea unor structuri secundare tip care sunt conservate remarcabil cu toate diferentele importante de secventa constate. Regiunile de secventa conservate sunt intotdeauna localizate la nivelul secventelor ne-imperecheate ale moleculei (Figura 4.14
Aceste molecule de ARN au conformatii globale tertiare complexe care iau nastere prin stivuiri multiple (asemanatoare celor din cazul ARNt) care compacteaza molecula in diferite domenii identificabile. De exemplu, moleculele de ARNr 16S cuprind patru domenii formate din motive secundare care poseda regiuni imperechiate scurte de mai putin de 10 nucleotide (Figura 4.14
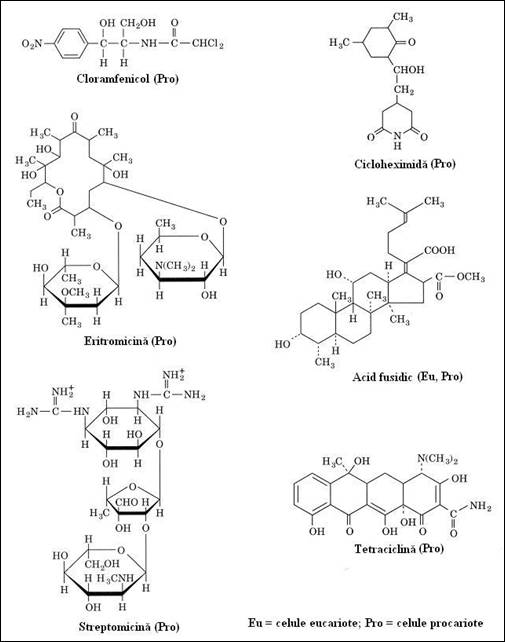
Figura 4. Diferite tipuri de antibiotice, majoritatea cu actiune selectiva asupra sintezei proteice la nivelul ribozomilor celulelor procariote
Rolul moleculelor ARNr
Desi nu putem afirma cu precizie ca se cunosc toate implicatiile functionale si structurale ale moleculelor de ARNr, se poate constata ca acestea: i) au rol structural intrand in compozitia ribozomilor; ii) faciliteaza fixarea celorlalte specii de ARN (ARNt si ARNm) la nivelul ribozomilor; iii) la procariote au rol de recunoastere, pe structura ARNm, a situsului de initiere a traducerii (AUG) prin hibridizarea unei secvente situata la capatul 3' al ARNr 16 S cu o secventa scurta, numita "Shine-Dalgarno", aflata la cateva nucleotide in amonte de AUG; iv) la eucariote nu se cunoaste inca cu precizie daca una dintre moleculele de ARNr recunoaste o secventa situata in apropierea AUG dar, se stie ca este absolut necesara recunoasterea extremitatii 5'"cap" a ARNm inainte de inceperea citirii informatiei acesteia.
ARN cu rol catalitic
Certitudinea ca numai proteinele au activitate enzimatica a fost total infirmata. Multa vreme identificarea enzimelor cu proteinele a condus la convingerea ca, numai proteinele, cu variatele lor structuri tridimensionale si grupari laterale, poseda flexibilitatea sa creeze situsuri active care sa catalizeze reactiile biochimice. Caracterizarea sistemelor implicate in procesarea ARN a demonstrat ca acest punct de vedere era deosebit de simplist.
T. Cech, in 1981 a facut o descoperire uimitoare studiind molecula ARNr 26S de la protozoarul termofil Tetrahymena. Acest ARN este derivat dintr-un precursor (ARN brut obtinut in urma transcrierii) care sufera un proces de maturare ("splicing") prin inlaturarea unui intron de 400 baze. Descoperirea importanta a constat in faptul ca reactia de maturare (inlaturarea intronului) este catalizata de insasi molecula de ARN. De altfel, intronul eliberat devine un catalizator activ in procesul de maturare a altor molecule de ARN.
In 1983, a fost descoperit un al doilea exemplu de cataliza realizata de o molecula de ARN. Sidney Altman si Norman Pace studiau in colaborare ribonucleaza P, enzima implicata in maturarea unui precursor ARNt la bacterii. Aceasta enzima are o structura neobisnuita fiind compusa dintr-o proteina si o molecula de ARN. De fapt, cercetarile efectuate au demonstrat ca activitatea enzimatica era datorata moleculei de ARN, partea proteica avand numai rol de structurare conformationala a ribonucleazei P. Spre deosebire de exemplul studiat de Cech, ARN din ribonucleaza P actiona asupra unei alte molecule substrat si nu asupra propriei structuri.
Numeroase tipuri de reactii catalitice sunt acum cunoscute ca fiind catalizate de ARN. Au fost denumite ribozime acele molecule de ARN dotate cu capacitate enzimatica despre care, pana la un anumit moment, se considera ca este rezervata numai proteinelor. Activitati catalitice ale ARN sunt directionate catre: i) substrate separate; ii) propria molecula si sunt descrise ca auto-maturare sau "auto-splicing", in functie de tipul de reactie.
Cateva din cele mai cunoscute exemple de activitate catalitica a ARN sunt:
a) Enzima ribonucleaza P, ribonucleoproteina care contine o singura molecula de ARN legata la o proteina. ARN poseda capacitatea de a cataliza clivarea substratului ARNt, in timp ce componenta proteica are un rol indirect, probabil in mentinerea structurii conformationale a ARN catalitic.
b) Intronii din grupa I poseda capacitatea de a-si realiza propria maturare din pre-ARNm in care sunt continuti. Reactia poate fi realizata in vitro numai de catre ARN, dar in vivo ea este asistata de proteine. Actiunea intronilor din grupa I genereaza molecule ARN care poseda multe alte activitati catalitice corelate cu activitatea de origine.
c) Anumiti introni, atat din grupul I cat si din II, contin faze de lectura care pot fi traduse in proteine. Unii introni din grupa I codifica pentru maturaze si pentru endonucleaze.
d) Moleculele mici de ARN din clasa viroizilor si virusoizilor au capacitatea de a realiza reactii de auto-clivare. Cu toate ca aceste reactii sunt intramoleculare, molecula poate fi impartita intr-o parte "enzimatica" si o parte "substrat" ale caror functii sunt independente. S-a demonstrat ca majoritatea viroizilor si virusoizilor care sufera auto-clivare au, in principiu, o structura secundara sub forma de "cap de ciocan" In figura 4.15 este prezentata aceasta structura obtinuta prin analize cu craze X. La nivelul structurii secundarea a complexului ARN-ADN sunt reprezentate prin linii punctate interactiile de tip Watson-Crick iar cele diferite prin linii continue.

Figura 4.14. Tipuri de structuri secundare posibile ale ARNr de tip 16S: a) la archeobacterii; b) eucariote, drojdii; c) mitocondrie de mamifer (bovine). Se observa conservarea filogenetica a domeniilor si subdomeniilor din structura (dupa Biochemistry, Voet&Voet, 1995)

Figura 4. Structura secundara "cap de ciocan a unei ribozime (Lewin B., Genes VII,2000)
Reactiile de inlaturarea a intronilor si sudare a exonilor ("splicing") sunt asistate de o serie de molecule de ARN mici " snRNA - small nuclear RNA". In procesul de "splicing" sunt implicate cinci molecule mici bogate in uracil (U1, U2, U4, U5, si U6) cu dimensiuni cuprinse intre 107 la 210 nucleotide. Asocierea secventiala a acestora cu un numar de sase la zece proteine formeaza "snRNP" ("small nuclear ribonucleoprotein particles") care impreuna cu pre-ARNm constituie complexul ribonucleoproteic numit spliceozom (Figura 4.16 si

Figura 4. . Asamblarea moleculelor de ARN U pentru formarea spliceozomului implicat in inlaturarea intronilor si sudarea exonilor din structura pre-ARNm (dupa Molecular Cell Biology Lodish H., et. al., 2000)
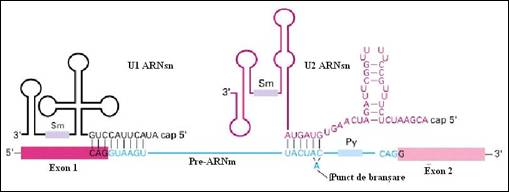
Figura 4. . Molecule ARNsn implicate in recunoasterea pre-ARNm si in realizarea procesului de "splicing" (dupa Molecular Cell Biology Lodish H., et. al., 2000)
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |