
Citogenetica umana moderna a aparut abia dupa 1956 cand Tjio si Levan au stabilit ca numarul corect, diploid de cromozomi ai speciei umane este de 46. Tot ei au fost aceia care au descris principalele caractere morfologice ale cromozomilor umani. Omul a cunoscut deci nucleul atomului inainte de a cunoaste numarul exact de cromozomi din celulele propriului sau corp. Aceasta intarziere a fost determinata de dificultati tehnice considerabile. Singurul material disponibil era reprezentat de sectiunile histologice taiate la o grosime care sa permita pastrarea intacta a garniturii de cromozomi dintr-o celula. Cromozomii erau aglomerati iar utilizarea fotografiei era imposibila datorita numeroaselor suprapuneri. In aceste conditii singura metoda de studiu a fost reprezentata de examenul microscopic in diferite planuri focale, urmat de redarea grafica prin desene a imaginilor.
Utilizand culturi de fibroblasti derivati din plamanii unor fetusi umani avortati, Tjio si Levan au observat in mod constant un numar de 46 de cromozomi. Corectitudinea acestor observatii a fost la scurt timp confirmata de Ford si Hammerton, care pe un material provenit din biopsii testiculare au numarat 46 de cromozomi in metafazele spermatogoniilor si 23 de bivalenti in spermatocitele primare aflate in metafaza I. Aceste cercetari, cat si cele care au urmat au beneficiat de o serie de imbunatatiri tehnice dintre care cele mai importante au fost:
utilizarea culturilor de tesuturi. Celulele cultivate in vitro prezinta un numar mai mare de mitoze iar calitatea lor este superioara deoarece agentii hipotonizanti si fixatori pot actiona mai bine asupra celulelor;
pretratarea celulelor cu colchicina (sau produsul sintetic similar denumit colcemid) blocheaza in mod specific celula in metafaza ca urmare a interactiunii cu fusul de diviziune. De asemenea, colchicina contracta cromozomii permitind o mai buna individualizare a acestora;
utilizarea solutiilor hipotone determina tumefierea celulelor si permite separarea cromozomilor dispusi intr-un spatiu mai mare;
strivirea celulelor (tehnica squash) determina dispozitia cromozomilor (separati datorita tratamentului hipotonic dar aflati inca intr-o celula de forma sferica) intr-un singur plan focal si permite astfel caracterizarea morfologica a cromozomilor.
Ca urmare a imbunatatirii tehnicilor de lucru a devenit posibila nu numai numararea cromozomilor ci si studierea trasaturilor morfologice ale fiecarui cromozom in parte. In felul acesta s-a putut stabili cauza sindroamelor de trisomie 21, 13 si 18 precum si a sindroamelor Turner si Klinefelter.
Introducerea metodei autoradiografiei a permis o identificare mai precisa a cromozomilor precum si identificarea unor modificari cromozomiale minore (deletii, inversii, translocatii) ce duceau la aparitia unor sindroame clinice corespunzatoare.
In anul 1970 au aparut tehnicile de bandare cromozomiala datorita lucrarilor de pionierat ale lui Cassperson si colab. care au demonstrat ca prin colorare cu quinacrina si examinare in UV se obtine o diferentiere longitudinala a cromozomilor sub forma unei succesiuni de benzi clare si intunecate. Modelul de bandare este caracteristic pentru fiecare pereche de cromozomi omologi in parte.
CROMOZOMII UMANI STRUCTURA SI NUMAR
Cromozomii sunt structuri filamentoase localizate in nucleu. Termenul de cromozom inseamna in stricto senso corpuscul colorat si deriva din limba greaca in care soma=corp iar chroma=culoare.
Cea mai mare parte a cunostintelor privitoare la structura cromozomilor umani a fost obtinuta prin studii de microscopie optica. Coloranti speciali, absorbti specific de ADN au permis identificarea fiecarui cromozom in parte. Cromozomii sunt mai usor de observat in timpul diviziunii celulare cand ating un maxim de condensare iar genele nu mai pot fi transcrise. Lungimea cromozomilor este variabila in cursul diviziunii, considerandu-se ca sunt mai lungi in profaza si anafaza decat in metafaza
Celulele tinere (inainte de faza de replicare a ADN) detin o singura macromolecula de ADN bicatenar, formand cromozomii monocromatidici. Dupa faza S de replicare a ADN, cromozomii devin bicromatidici, posedand doua macromolecule de ADN bicatenar. Cele doua cromatide ale unui cromozom dispuse aproape paralel, sunt reunite in regiunea centromerului, regiune denumita si constrictie primara. Prin centromer se intelege o regiune distincta a cromozomului prin care acesta se ataseaza in mod reversibil de microtubulii fusului de diviziune, fapt ce-i permite sa se orienteze si sa migreze spre unul din cei doi poli ai celulei in timpul anafazei. Deoarece centromerul determina deplasarea cromozomilor el a mai fost denumit si kinetochor. Centromerul imparte cromozomul in doua: bratul lung (q) si bratul scurt (p) si are o pozitie variabila dar constanta pentru acelasi cromozom. In functie de pozitia centromerului cromozomii pot fi: metacentrici cand centromerul este localizat median iar cele doua brate sunt de lungimi aproape egale; submetacentrici cand centromerul este plasat inafara regiunii mediane iar unul din brate este mai lung decat celalalt si acrocentrici cand centromerul este plasat foarte aproape de unul din capete si cromozomul apare format din doua brate inegale dintre care unul foarte lung si altul foarte scurt.
Unii cromozomi prezinta pe langa constrictia primara si o constrictie secundara care izoleaza de restul cromozomului o portiune denumita satelit sau trabant.
Extremitatea fiecarui cromozom poarta numele de telomer. Telomerele joaca un rol esential in mentinerea stabilitatii si integritatii cromozomilor. La om, acestea sunt formate din mai multe secvente TTAGGG, repetate in tandem, care sunt mentinute de enzime denumite telomeraze. Se apreciaza ca reducerea nivelului telomerazelor asociata cu scaderea numarului de secvente TTAGGG joaca un rol important in procesele de imbatranire si moarte celulara.
Nucleul fiecarei celule somatice contine doua seturi de cromozomi, fiecare mostenit de la un parinte. Membrii unei perechi de cromozomi sunt numiti cromozomi omologi.
Materialul din care sunt formati cromozomii, ADN si proteine histonice, poarta numele de cromatina.
Gradul de condensare colorare a cromozomilor nu se realizeaza uniform. Acest lucru se observa mai ales in profaza si teleofaza. Ca urmare se apreciaza ca cromatina prezinta doua stari functionale alternative si reversibile: eucromatina si heterocromatina. Ele rezulta din faptul ca cromatina poate avea grade de condensare diferite in diferite regiuni ale cromozomilor. Gradul de condensare al fibrei de cromatina este corelat cu activitatea transcriptionala, cromatina nefiind exprimata cand se gaseste in stare de condensare maxima. Cromozomii mitotici reprezinta un caz extrem, in sensul ca ei sunt transcriptional inerti, in celula transcriptia nerealizandu-se ii timpul diviziunii celulare. Eucromatina reprezinta zonele mai slab condensate ale cromatinei, zone care se coloreaza mai putin intens cu coloranti specifici. La nivelul ei se gaseste cea mai mare parte a proteinelor nonhistonice si secventele unice de ADN; ca urmare reprezinta partea activa genetic a cromatinei interfazice. Heterocromatina reprezinta acele regiuni ale genomului care sunt ii interfaza intr-o stare de intensa condensare si nu se exprima adica este inactiva din punct de vedere transcriptional. Condensarea materialului genetic este probabil responsabila de inactivitatea sa. Trebuie sa precizam insa ca reversul acestei afirmatii nu este adevarat in sensul ca genele active se gasesc intr-adevar la nivelul eucromatinei, dar numai o mica parte din secventele eucromatice sunt transcrise. Localizarea in eucromatina este deci o conditie necesara dar insuficienta pentru exprimarea unei gene.
La om, nucleul celulelor normale contine 46 de cromozomi impartiti in perechi, existand astfel 22 de perechi de autosomi si o singura pereche de cromozomi de sex XX la femeie si XY la barbat. Celulele somatice sunt denumite diploide continand toti cei 46 de cromozomi in timp ce gametii (ovulul si spermatozoidul) sunt haploizi continand doar 23 de cromozomi.
Numarul total de cromozomi la diferite organisme variaza considerabil dar este constant pentru o anumita specie. La unele specii de feriga nucleul contine 500 de cromozomi in timp ce la o specie de crustacee se estimeaza ca in nucleu se gasesc intre 1500 si 1600 de cromozomi. La cealalta extrema se gaseste un vierme intestinal la care nucleul contine doar doi cromozomi. Unele maimute au, ca si omul, 46 de cromozomi. Primatele superioare insa, precum cimpanzeul, gorila si urangutanul au 48 de cromozomi. Dintre aceste primate cromozomii cimpanzeului seamanna cel mai bine cu cei umani, existand o diferenta de doar 1%. Se considera ca cromozomul nr. 2 de la om ar fi rezultatul unei fuziuni a doi cromozomi de la cimpanzeu, celelalte diferente intre complementul cromozomial de la cele doua specii fiind datorate numeroaselor inversii peri- si paracentrice. Aceste observatii au fost exploatate de specialistii in biologie moleculara pentru cartarea si clonarea genelor umane.
Cromozomii X si Y sunt cunoscuti sub denumirea de cromozomi ai sexului datorita rolului lor crucial in determinarea sexelor. La om precum si la majoritatea mamiferelor atat femela cat si masculul au doi cromozomi de sex- XX la femela si XY la mascul. Cromozomul Y este mult mai mic decat X si poarta doar cateva gene de importanta functionala, cea mai importanta fiind SRY sau factorul de determinare testiculara.
La femeie fiecare ovul poarta cate un cromozom X, pe cand la barbat spermatozoidul poate purta ori X ori Y. Sansa ca un spermatozoid ce poarta cromozomul X sau Y sa fecundeze un ovul este aproximativ egala, asa cum si numarul de produsi de conceptie de sex masculin si feminin este aproximativ egal. In realitate, se nasc mai multi copii de sex masculin ca si cei de sex feminin desi in timpul copilariei si maturitatii sex ratio-ul ajunge 1:1.
METODE DE ANALIZA CROMOZOMIALA
Cu unele imbunatatiri, metodele folosite de Tjio si Levan sunt folosite si astazi in aproape toate laboratoarele de citogenetica in scopul realizarii unei analize a constitutiei cromozomiale a unui individ. In acest sens se realizeaza cariotipul, adica aranjarea sistematica standardizata a cromozomilor unei celule, fotografiati la microscop in timpul metafazei mitozei.
PREPARAREA CROMOZOMILOR
Pentru a studia cromozomii umani pot fi folosite practic orice tesuturi cu celule vii, nucleate care se pot divide. Cel mai frecvent sunt folosite limfocitele din sangele periferic desi probe pentru analiza cromozomiala pot fi preparate relativ usor folosind piele, maduva hematogena, vili corionici sau amniocite( celule provenite din lichidul amniotic).
In cazul in care pentru obtinerea preparatelor cromozomiale se foloseste sangele periferic (venos), limfocitele sunt mai intai separate si trecute apoi pe un mediu de cultura la care se adauga fitohemaglutinina care stimuleaza limfocitele T sa se divida. Celulele sunt tinute in conditii sterile la 37oC aproximativ 3 zile, timp in care ele se vor divide. Inainte de realizarea preparatelor se adauga colchicina. In final celulele sunt separate din cultura si tratate cu solutie hipotona care produce umflarea celulelor si dispersia cromozomilor. Preparatele sunt apoi fixate pe o lama de microscop iar dupa colorare vor putea servi analizei citogenetice.
Tehnicile conventionale asigura o colorare uniforma, nebandata a cromozomilor. Ele sunt la ora actuala mai putin folosite datorita apaitiei posibilitatii de bandare cromozomiala. Bazele chimice ale reactiilor de colorare sunt inca insuficient cunoscute.
Prin tehnicile de bandare de-a lungul cromozomilor apare o succesiune de benzi transversale. Aceasta alternanta de benzi clare cu benzi intunecate este identica la cromozomii omologi si diferita pentru cromozomii apartinand diferitelor perechi de cromozomi omologi.
Bandarea Giemsa
Bandarea Giemsa a devenit astazi una dintre cele mai cunoscute si utilizate tehnici de bandare pentru cromozomii mamiferelor. Cele mai uzuale metode pentru a obtine aceasta colorare se bazeaza pe tratarea lamelor cu preparate cromozomiale cu o proteaza (de regula tripsina) sau pe incubarea lamelor in solutie citrat salina fierbinte, desi exista o mare varietate de alte metode. Amprenta cromozomului bandat se apreciaza ca reflecta atat compozitia sa structurala, cat si cea functionala. Benzile inchise la culoare, corelate cu cromomerele din pachiten, isi replica ADN-ul in faza S tarzie, si contin portiuni bogate in T-A. Se pare ca ele contin putine gene active si pot diferi de benzile deschise la culoare prin proteinele cromozomiale. Extractia diferentiata a proteinelor in pretratamentul de fixare si bandare este importanta in mecanismul de obtinere a benzilor G.
Bandarea Q
Quinacrina este un colorant acridinic care se leaga de ADN fie de ionii externi, fie prin intercalare. Bandarea in acest sistem pare a fi puternic influentata de variatia in compozitia proteica a cromozomilor. Bandarea cu quinacrina duce la aparitia benzilor Q care sunt asemanatoare ca dimensiuni si dispozitie benzilor G. Acest tip de bandare permite insa o mai buna evidentiere si analiza a unor portiuni cromozomiale de mare importanta precum regiunile satelitare ale cromozomilor acrocentrici si regiunile centromerice ale cromozomilor 1, 9 si 16 . Polimorfismul acestor regiuni, ca si variatiile cromozomului Y, pot fi mai usor observate cu un astfel de sistem de bandare.
Bandarea cu quinacrina este relativ simpla, rapida si nu necesita pretratamente speciale inainte de colorare. Metoda da rezultate bune chiar si cu lame mai vechi sau chiar in cazul unor preparate cromozomiale mai putin reusite.
Bandarea de reversie
Benzile care apar ca deschise la culoare prin bandarea Giemsa, vor aparea ca avand o coloratie intensa prin bandare de reversie. Benzile R pot fi obtinute prin incubare in solutie salina fierbinte inainte de colorare Giemsa, sau prin colorare cu acridin oranj. O alta metoda implica colorarea cromozomilor cu cromomicina A (un colorant fluorescent), urmata de o a doua colorare cu verde de metil care se leaga de perechile de baze azotate complementare celor de care se leaga cromomicina A. Ca urmare, vor ramane fluorescente doar regiunile in care perechile de baze sunt predominant de un singur tip.Studiul in microscopia electronica de tip scanning a cromozomilor umani bandati G releva in cromatidele cromozomilor regiuni proeminente care alterneaza cu santuri circulare.Santurile circulare corespund benzilor G pozitive iar regiunile proeminente corespund benzilor R pozitive observate in microscopia optica.
Bandarea C (heterocromatina centromerica)
Acest tip de bandare se obtine dupa tratamente de denaturaresi renaturare ale cromozomilor urmate de colorare cu solutie Giemsa. Se obtine astfel o coloratie intensa a heterocromatinei constitutive din regiunile pericentromerice care sunt caracteristice in ce priveste intinderea pentru fiecare cromozom in parte. Cromozomii 1,9 si 16 contin cate un segment mare de heterocromatina extins in bratul q. romosomul romosomul Y prezinta un segment heterocromatic in jumatatea distala a bratului q ce corespunde segmentului q stralucitor si o banda ingusta la nivelui centromerului.Satelitii au o coloratie mai fina decat regiunile centromerice.Bandarea C permite evidentierea variatiilor individuale sau familiale ale constrictiilor secundare si ale heterocromatinei pericentromerice precum si detectarea centromerilor in cazurile de remanieri cromozomiale. Aparitia ocazionala de benzi C cu localizare terminala sau intercalara are la origine dislocarea heterocromatinei din regiunea centromerica prin inversii sau translocatii.
Bandarea T
Benzile T (T=telomere) se obtin in urma unei denaturari termice mai pronuntate necesara pentru suprimarea benzilor R de pe bratele cromozomilor cu exceptia telomerelor. Aceste benzi reprezinta fractiunea cea mai rezistenta la tratamentul termic al benzilor R. Dupa colorare cu acridin orange cromozomii observati in lumina UV prezinta o coloratie slaba portocalie iar telomerele au o fluorescenta verde a carei intensitate variabila permite diferentierea cromozomilor intre ei. Acest tip de mercaj este util intrucat permite depistarea unor modificari minore ce vizeaza extremitatile cromozomiale.
Bandarea de rezolutie inalta
Bandarea G permite in mod normal identificarea a circa 400 de benzi per set haploid de cromozomi umani. Fiecare dintre aceste benzi corespunde la circa 8000 kb de ADN. Bandarea de rezolutie inalta a cromozomilor aflati in profaza sau prometafaza (cand au un nivel de condensare mai scazut) permite evidentierea a circa 800 de benzi per set haploid de cromozomi. Aceasta tehnica necesita inhibarea diviziunii celulare cu un antagonist al acidului folic precum metotrexatul. Acidul folic este adaugat mediului de cultura pentru a permite intrarea celulelor in mitoza.Ulterior este adaugata colchicina la un anumit interval de timp, si anume atunci cand cea mai mare parte din celule vor fi in prometafaza.
ANALIZA CROMOZOMIALA
In vederea realizarii cariotipului cromozomii trebuie mai intai identificati cu precizie si aranjati in perechi. Caracterizarea morfologica a cromosmilor metafazici are la baza o serie de criterii stabilite de ISCN-International System for Human Cytogenetic Nomenclature.
1.Lungimea relativa a unui cromozom este reprezentata de raportul dintre
lungimea cromozomului( reprezentata de suma lungimii bratelor p+q) si lungimea totala a unui set haploid de cromozomi(22 de autosomi +X).
Lungimea absoluta a cromozomilor nu are o importanta prea mare pentru caracterizarea acestora deoarece variaza in functie de stadiul metafazei, cromozomii fiind mai lungi in metafaza precoce (prometafaza) decat in metafaza tardiva. Agentii statmochinetici utilizati in mod curent in citogenetica pentru blocarea mitozelor in metafaza (colchicina, colcemid, vinblastina) accentueaza aceasta scurtare.
2. Indicele centromeric este definit prin raportul dintre bratul scurt (p) si lungimea totala a cromozomului.
IC=p/(p+q)
3. Raportul bratelor este reprezentat de dimensiunea bratului lung (q) raportata la dimensiunea bratului scurt (p).
B=q/p
La cromozomii metacentrici raportul este de 1.0-1.7, la cei submetacentrici 1.7-3.0, la subtelocentrici 3.0-7.0 iar la cei acrocentrici 7.0-4
4.Existenta satelitilor. Satelitii sunt prezenti la cromozomii acrocentrici si sunt reprezentati de mase mici de cromatina. Ei se gasesc la extremitatea terminala a bratelor scurte a cromozomilor de care sunt separati prin constrictii secundare.
5.Constrictii secundare. Termenul de constrictie primara poate fi considerat sinonim cu cel de centromer. Unii cromozomi prezinta insa constrictii si in alte regiuni ale bratelor, denumite constrictii secundare.
6.Replicarea asincrona a cromozomilor. Evidentierea replicarii asincrone a cromozomilor umani in cadrul perioadei S a ciclului celular a fost posibila in urma administrarii de H3 timidina si urmarirea incorporarii acesteia in cromozomi in functie de momentul replicarii. Metoda a permis deferentierea cromozomilor din grupele B,D si E. De asemenea s-a pus in evidenta replicarea tardiva a unuia din cei doi cromozomi X la femei.
7.Modelul de benzi cromozomiale se refera la obtinerea unor coloratii diferentiale ale cromozomilor.
Pe baza criteriilor mentionate, cromozomii umani au fost repartizati in 7 grupe morfologice.
Grupa A (1-3). Cuprinde cromozomi mari (9,08-7,06F) dintre care 1 si 3 sunt metacentrici iar cei din perechea 2 sunt submetacentrici. Cromozomii din perechea 1 prezinta o constrictie secundara in regiunea proximala a bratului q.
Grupa B (4-5). este alcatuita din cromozomi lungi submetacentrici si au dimensiuni situate intre 6.55-6,13F
Grupa C (6-12) si X cuprinde cromozomi de marime mijlocie (5,84-4,46F). Perechile 6,7,8,11 au centromerul situat in regiunea mediana (sunt metacentrici) iar perechile 9,10 si 12 au centromerul situat submedian deci sunt submetacentrici. Cromozomii din perechea 9 prezinta o constrictie secundara in regiunea proximala a bratului q. Cromozomii X sunt metacentrici si au dimensiuni similare cu a cromozomilor din perechile 7 si 8.
Grupa D (13-15) cuprinde cromozomi de marime mijlocie (3,64-3,36F) acrocentrici. Toti cromozomii acestei grupe prezinta sateliti.
Grupa E (16-18) cuprinde cromozomi relativ mici (3,23-2,76F) cu centromerul situat median la cei din perechea 16 si submedian la perechile 17 si 18.Cromozomii din perechea 16 au o constrictie secundara in regiunea proximala a bratului q.
Grupa F (19-20) cuprinde doua perechi de cromozomi mici (2,52-2,33F), metacentrici.
Grupa G (21-22) cuprinde cei mai mici cromozomi ai cariotipului uman (1,83-1,68F), sunt acrocentrici si prezinta sateliti.
Cromozomul Y are lungimea apropiata de cea a cromozomilor din grupa G (1.96F), este de tip acrocentric dar spre deosebire de ceilalti cromozomi acrocentrici din cariotip nu au sateliti.
HIBRIDIZARE FLUORESCENTA IN SITU (FISH)
Metodele noi de diagnostic combina tehnicile citogenetice conventionale cu cele de inginerie genetica moleculara. Tehnica FISH se bazeaza pe capacitatea unei portiuni de ADN monocatenar de a se lega de o secventa complementara care poate fi localizata oriunde pe o placa metafazica. Spre deosebire de alte metode folosite in analiza cromozomilor tehnica FISH poate sa fie aplicata si celulelor aflate in interfaza.
In aceasta tehnica proba de ADN este conjugata cu nucleotide modificate care dupa hibridizare cu ADN provenit de la pacient permite regiunii unde a avut loc hibridizarea sa devina vizibila in lumina UV. Aceasta tehnica este larg folosita astazi in scopul diagnosticarii clinice si datorita faptului ca rezultatele se obtin foarte rapid.
POLIMORFISMUL CROMOZOMILOR UMANI
Studiile au aratat ca anumite regiuni ale cromozomilor prezinta variatii ale dimensiunii sau ale afinitatii fata de coloranti. Intrucat acest polimorfism nu are o expresie fenotipica deci nu exercita nici o influenta asupra fenotipului, diferentele structurale mentionate sunt considerate variante normale.
Variabilitatea cromozomiala implica regiunile constrictiilor secundare, portiunea distala a cromozomului Y si satelitii. Benzile C si Q pot prezenta variatii ale lungimii in cadrul diferitelor grupe de cromozomi, fiind caracteristice pentru fiecare individ in parte.Benzile mai mari rezulta din repetarea in tandem a unor unitati mai mici, identice. Cromozomii 1, 9, 16 au benzi care variaza atat in lungime cat si in localizarea lor in regiunea centromerica si a constrictiilor secundare. In majoritatea cazurilor banda este extinsa in bratul lung si foarte rar in bratul scurt ca rezultat al unor inversii pericentrice. Variatia in lungime a benzilor C in acesti cromozomi se datoreaza in primul rand variatiei in lungime a constrictiilor secundare.
Cromozomul Y are o banda heterocromatica foarte ingusta la nivelul centromerului, pentru care diferentele de lungime de la un cromozom la altul sunt dificil de raportat. Segmentul distal al bratului q poate insa varia considerabil in lungime.
Au fost semnalate urmatoarele tipuri de variante cromozomiale:
Constrictie secundara alungita la cromozomii 1, 9, 16. Se considera constrictia secundara ca fiind alungita atunci cand are o lungime cel putin dubla fata de lungimea constrictiei secundare a cromozomului omolog.
Bratul p alungit la cromozomii acrocentrici. Bratul p se considera alungit atunci cand este egal sau mai lung decat bratul scurt al unui cromozom din perechea 18.
Sateliti proeminenti. Sunt definiti astfel cand sunt mai mari decat bratul scurt de la acelasi cromozom.
Sateliti dubli- se remarca prin colorare doi sateliti in tandem.
Cromozom y mare sau mic. Cromozomul Y este considerat mare atunci cand Y/F>1,00 si mic atunci cand raportul este de sub 0,7.
Prezenta satelitilor pe bratul scurt al cromozomului 17 (17ps). Aceasta varianta se intalneste foarte rar, semnalandu-se in cateva cazuri printre pacientii internati in clinicile de psihiatrie.
Prezenta satelitilor pe bratul q al cromozomului y. Este o varianta foarte rar intalnita fiind semnalata fie la pacienti normali fie la pacienti cu sindromul Klinefelter.Acest caz particular se pare ca rezulta in urma translocatiei satelitilor unui cromozom din grupele D sau G pe bratul q al cromozomului Y.
ANOMALII (ABERATII CROMOZOMIALE)
O mare parte a maladiilor conditionate genetic este datorata aberatiilor (mutatiilor) cromozomiale numerice sau structurale.
Aberatii numerice
Aberatiile numerice pot fi impartite in doua mari categorii si anume:
1. aneuploidia - datorata pierderii sau dobandirii unuor cromozomi suplimentari;
2. poliploidia datorata aditiei unuia sau a doua seturi haploide complete
1. Aneuoloidia
Cele mai frecvent intalnite aneuploidii sunt trisomiile datorate aparitiei unui cromozom suplimentar, tetrasomiile datorate existentei in celule a doi cromozomi suplimentari pentru o anumita pereche de cromozomi omologi si monosomiile rezultate ca urmare a pierderii unui cromozom.
Sindromul Down este datorat unui cromozom suplimentar din perechea 21 si ca urmare este cunoscut si sub numele de trisomie 21. Exista si alte trisomii ale autosomilor compatibile cu supravietuirea precum sindromul Patau (trisomia 13) si sindromul Edwards (trisomia 18). Cele mai multe trisomii ale autosomilor duc insa la pierderea timpurie a sarcinii, trisomia 16 fiind relativ frecvent intalnita in cazul avorturilor spontane in primul trimestru al sarcinii. Prezenta in exces a unui heterosom (X sau Y) are efecte fenotipice mai putin grave.
Originea aneuploidiei
Principalele cauze ale aneuploidiei sunt: non-disjunctia, pierderea de cromozomi datorita retardarii anafazice, endoreduplicarea selectiva si mitozele multipolare.
Non-disjunctia este procesul prin care cromozomii omologi (sau cromatidele aceluiasi cromozom) nu se separa pentru a se distribui in mod egal celulelor fiice, ci migreaza impreuna in una din aceste celule. Acest proces poate avea loc atat in timpul gametogenezei cat si in primele stadii ale clivarii zigotului.
Procesul de non-disjunctie poate aparea in meioza I, meioza II sau (mai rar) in ambele diviziuni meiotice. Vorbim despre non-disjunctie primara cand celulele germinale sunt normale si non-disjunctie secundara ce opereaza pe celule germinale anormale (de ex. 47, xxx).
Consecintele non-disjunctiei sunt diferite la barbati si la femei, deosebire determinata de particularitatile gametogenezei la cele doua sexe. Astfel la barbat dintr-un spermatocit primar rezulta 4 spermatozoizi functionali; la femeie dintr-un ovocit primar rezulta un singur ovul functional, fiecare diviziune meiotica fiind insotita de eliminarea unui globul polar. Din acest motiv la barbati non-disjunctia in decursul meiozei I va determina aparitia unor spermatozoizi disomici si nulisomici in timp ce non-disjunctia in meioza II va produce si gameti normali alaturi de gameti disomici si nulisomici.. La femeie insa non-disjunctia survenita fie in decursul primei, fie a celei de-a doua meioze nu poate produce decat ovule anormale: disomice sau nulisomice. Posibilitatea ca un zigot anormal sa fie consecinta unei anomalii survenite in timpul ovogenezei este deci mai mare decat cea a unei anomalii similare aparute in decursul spermatogenezei. Studiile au aratat ca cea mai mare parte a copiilor cu trisomii autosomaleau mostenit cromozomul suplimentar de la mama (Tab. )
Tabelul nr. Originea cromozomuli suplimentar in cazul celor mai frecvente trisomii
|
Tipul aberatiei cromozomiale |
Origine paterna (%) |
Origine materna (%) |
|
Trisomia 13 |
15 |
85 |
|
Trisomia 18 |
10 |
90 |
|
Trisomia 21 |
5 |
95 |
|
45,XXX |
5 |
95 |
|
47,XXY |
45 |
55 |
|
47,XYY |
100 |
0 |
|
|
|
|
Cu o frecventa redusa, non-disjunctia poate sa apara si in primele diviziuni mitotice ale zigotului. Vor rezulta doua sau mai multe linii celulare diferite, fenomen denumit mozaicism. Constitutia cromozomiala a organismului care rezulta depinde de momentul in care are loc non-disjunctia. Astfel, daca non-disjunctia are loc in timpul primei diviziuni de clivare a unui zigot normal, va rezulta un produs de conceptie care va prezenta doua linii celulare anormale, una monosomica si una trisomica pentru cromozomul in cauza. Daca non-disjunctia apare in a doua diviziune a zigotului, va rezulta un blastocit mixoploid alcatuit din trei linii celulare: una normala disomica, una trisomica si una monosomica. In cursul vietii indivizilor se poate produce o selectie care va duce la eliminarea celulelor dezavantajate (in special a celor monosomice). Intrucat aceasta selectie opereaza mai eficient in tesuturile cu diviziune rapida, proportia celulelor anormale nu va fi identica in toate tesuturile unei anumite persoane.
Cauzele non-disjunctiei
Cauzele non-disjunctiei nu sunt inca suficient de bine cunoscute. Evenimentele non-disjunctionale pot aparea in decursul ambelor diviziuni meiotice ale spermatogenezei si ovogenezei, dar survin cu o frecventa mai mare in ovogeneza. In acest sens pledeaza faptul ca varsta medie a mamelor care au dat nastere unor copii ci trisomii 21, 13 sau 18, este mai ridicata decat a mamelor care au dat nastere unor copii normali (tab. de la S.Down). Acest fenomen ar putea fi explicat prin faptul ca spermatogeneza are loc continuu, in tot cursul vietii, in timp ce la femeie prima diviziune meiotica are loc inainte de nastere, ovocitele ramanand in ovar in stare de repaus pana in momentul ovulatiei care are loc intre 15-50 de ani. In tot acest interval de timp ovocitele sunt supuse unui proces de imbatrinire si unor influente externe (iradieri, infectii, medicamente) care pot reduce eficienta meiozei. Studiile au aratat, de asemenea, ca absenta recombinarii in profaza meiozei I ar predispune la non-disjunctie. Aceasta observatie nu este surprinzatoare deoarece chiasmata sunt responsabile de mentinerea conjugata a celor doi cromozomi omologi pana in diakineza cand se vor separa. La femeie recombinarea se produce inainte de nastere in timp ce non-disjuctia se produce cu 15-50 de ani mai tarziu. Aceasta observatie sugereaza faptul ca non-disjunctia ar putea fi cauzata de cel putin doi factori, si anume: absenta recombinarii intre cromozomii omologi in perioada fetala si aparitia, mult mai tarziu, unor anomalii in formarea fusului de diviziune.
O alta explicatie a aocierii intre varsta avansata a mamei si riscul nasterii unui copil afectat de o trisomie autosomala, ar consta in aceea ca supravietuirea embrionilor trisomici ar putea fi rezultatul unei diminuari a raspunsului imun al mamei, odata cu varsta. Se apreciaza, de asemenea, ca non-disjunctia ar fi favorizata si de cresterea intervalului de timp dintre ovulatie si fertilizare.
La unele specii precum cele ale genului Drosophila, s-a constatat ca non-disjunctia este controlata genetic. Aceasta observatie ar putea furniza o explicatie pentru familiile care par a fi predispuse la non-disjunctii repetate.
Procesul de retardare anafazica reprezinta o alta cauza a pierderii de cromozomi. In decursul anafazei, unul sau cativa cromozomi nu se deplaseaza spre polii celulei astfel incat dupa ce membrana celulara se reface in telofaza ei sunt exclusi din nucleu, pierzandu-se in citoplasma.
Endoreduplicarea selectiva a fost definita ca o replicare dubla a unei parti din genom (un cromozom intreg sau o parte de cromozom).
Mitozele multipolare sunt o cauza a anuploidiilor In special in celulele canceroase. Ele sunt consecinta formarii unui fus multipolar datorat unor diviziuni succesive ale centrosomului si duc la repartizari inegale ale cromozomilor in celulele fiice.
Un procent relativ mic din numarul de aneuploizi ar putea fi atribuit si altor factori precum:
tulburarilor autoimune parentale (anticorpi anttiroidieni la mama);
actiunea mutagena a iradierilor medicale;
infectiile virale materne.
2. Poliploidia
Organismele poliploide contin in celule un multiplu al numarului haploid de cromozomi. La om, formele triploide contin 69 de cromozomi in celule si au fost intalnite in special in cazul analizelor citogenetice a produsilor de conceptie avortati. Literatura de specialitate descrie doar cateva cazuri de nou-nascuti triploizi, dar acestia au murit la scurt timp dupa nastere. Zigotii triploizi pot lua nastere fie prin fecundarea unui ovul de catre doi spermatozoizi (dispermie), fie, prin fuziunea ovulului cu un globul polar (diginie). S-a constatat ca in cazul triploizilor rezultati prin dispermie placenta este marita, hidatidiforma in ce in cazul celor rezultati in urma dispermiei placenta este mai mica decat in situatii normale.
Tetraploizii au la om 92 de cromozomi si apar cu o frecventa foarte mica, tetraploidia fiind cosiderata incompatibila cu viata. Astfel de constitutii genetice au fost identificate in urma realizarii analizei cromozomiale a unor produsi de conceptie avortati spontan. Se apreciaza ca aceste forme apar, cel mai probabil, ca urmare a unei supresii a primei diviziuni de clivare a unui zigot diploid, dupa separarea cromozomilor dar inaite de diviziunea citoplasmei.
Poliploidizarea poate aparea si in decursul vietii adulte datorita unor accidente mitotice (endomitoza sau endoreduplicarea). Celule poliploide au fost evidentiate in proportie foarte redusa in maduva osoasa si in culturile de limfocite, fiind insa foarte frecvente in celulele tumorale. In cazul endomitozei cromozomii se comporta normal pana in profaza tarzie cand membrana nucleara nu se dezintegreaza iar fusul de diviziune nu se formeaza. Endoreduplicarea se caracterizeaza prin doua duplicari succesive a cromozomilor in interfaza astfel ca in profaza si metafaza se observa un numar dublu de cromozomi. Procesul de separare a cromozomilor se desfasoara in mod normal dar el opereaza pe o garnitura cromozomiala dubla si va duce in final la aparitia a doua celule tetraploide.
Aberatii structurale
Aberatiile structurale ale cromozomilor rezulta ca urmare a aparitiei unor rupturi urmate fie de pierderea segmentului lipsit de centromer, fie de realipirea fragmentului rupt intr-o configuratie diferita.
Aberatiile cromozomiale pot fi balansate sau nebalansate. In cazul celor balansate complementul cromozomial ramane complet deoarece nu a avut loc o pierdere sau un castig de material genetic sau nebalansate. Ca urmare acest tip de aberatii cromozomiale nu are in general urmari mai putin rarele situatii in care unul din punctele de ruptura este localizat la nivelul unei gene functionale importante. Purtatorii unor aberatii cromozomiale balansate prezinta un risc de a avea copii cu un complement cromozomial anormal. In cazul aberatiilor cromozomiale nebalansate, se produce o modificare a cantitatii de material genetic iar urmarile asupra organismului sunt cel mai adesea severe.
Rupturile
Constau in intreruperi ale continuitatii cromatidelor sau cromozomilor insotite de o deplasare laterala a fragmentelor distale. Ele pot avea trei consecinte:
reunirea fragmentelor cu refacerea structurii initiale a cromozomilor (restitutie);
reunirea cu modificare rezultand diferite tipuri de aberatii cromozomiale;
lipsa reunirii cu pierderea fragmentului rupt.
Agentii care provoaca rupturi cromozomiale (agenti clastogeni) sunt destul de variati. Ei pot fii clasificati in factori externi (radiatii, substante chimice, virusuri) si interni (afectiuni ereditare, imbatranirea). Expunerile la doze mari de radiatii pot fi accidentale sau datorate iradierilor profesionale sau medicale. In grupa agentilor chimici intra substante extrem de numeroase utilizate pe scara larga (chimioterapicele antitumorale, unele antibiotice, pesticide, detergenti si aditivi din industria cosmetica sau alimentara). Consecintele actiunii virusurilor infectioase (virusurile hepatitei A si B, rubeolei) sau oncogene (polyoma, SV-40) sunt extrem de grave iar tabloul modificarilor cromozomiale este complex. In unele afectiuni ereditare precum Sindromul Bloom sau anemia Fanconi exista in mod spontan un nivel crescut de rupturi cromozomiale. Persoanele care sufera de aceste boli prezinta o incidenta crescuta a anumitor tipuri de cancer.
Analizand rupturile aparute spontan in limfocitele umane s-a inregistrat un mare numar de puncte de ruptura care au fost catalogate in functie de localizarea lor in benzile negative sau pozitive. Din studiile amintite mai sus se desprind urmatoarele aspecte:
rupturile spontane ca si cele induse sunt localizate de preferinta in benzile G sau Q negative;
rupturile dintr-un tip de banda tind sa se reuneasca cu cele apartnand aceluiasi tip de banda (negativ-negativ, pozitiv-pozitiv), combinatiile heterologe fiind rar intalnite;
frecventa rupturilor este mai mica in regiunile centromerice si telomerice;
distributia rupturilor nu este proportionala cu lungimea cromozomilor. Cel mai mare numar de rupturi pe unitatea de lungime o prezinta in ordine cromozomii 21, Y, 18, 9 si 13, iar cel mai mic numar de rupturi s-a observat la cromozomii 16, 6, 2, 3, 19 si 20.
Deletiile
Deletiile (lipsurile) constau in piederea unui fragment de cromatida sau de cromozom. Ele pot fi terminale sau interstitiale si sunt cauzate de rupturi care apar la nivelul cromozomului. Daca ruptura a avut loc inainte de faza S a ciclului celular va genera o deletie cromozomiala, iar dupa aceasta faza o deletie cromatidica. In urma unor deletii izocromatidice suprafetele de ruptura de la capetele cromatidelor pot fuziona, iar in faza urmatoare a diviziunii, cand cei doi centromeri rezultati prin clivare migreaza spre cei doi poli ai fusului de diviziune, cromatidele raman unite prin zona de fuzionare realizand o punte anafazica. In anafaza, fragmentele telomerice acentrice nu se pot deplasa spre unul dintre polii celulei datorita faptului ca sunt lipsite de centromer. Din aceasta cauza se pierd si rezulta o lipsa terminala simpla. Daca se produc rupturi simultane la ambele capete ale unui cromozom, zonele acentrice se pierd, iar fragmentul cu centromeri formeaza un cromozom circular (inelar) prin unirea capetelor lipsite de telomere care devin instabile si au tendinta de a se uni.
Deletiile sunt datorate, cel mai frecvent, formarii unei bucle de incalecare, la nivelul careia se produce o ruptura dubla rezultand patru capete. Va rezulta un fragment acentric (care la nivelul metafazei formeaza micronucleii) si un cromozom cu deletie. Contrar asteptarilor deletiile intercalare sunt mai frecvente decat cele terminale, deoarece nu implica formarea de noi telomere pentru stabilizarea deletiilor.
Deletia consta in pierderea un fragment de cromozom si prin urmare va duce la aparitia unei monosomii pentru fragmentul pierdut. In general o deletie mare (mai mare de 2%dintr-un set haploid de cromozomi) este considerata incompatibila cu supravietuirea.
Unele deletii, precum cele care duc la aparitia sindroamelor Wolf-Hirschhorn sau cri-du chat pot fi observate la microscop in preparate obtinute din placi metafazice. In cazul in care deletia implica un fragment de dimensiuni foarte mici identificarea se poate face numai prin realizarea unei analize citogenetice in prometafaza si studiilor de hibridizare in situ. Asa este cazul sindroamelor Angelman si Prader-Wili.
Duplicatiile

Duplicatia consta in dublarea unui fragment cromozomial sau
cromatidic.
Fig. nr.
Aparitia unei duplicatii este consecinta unei rupturi urmate de anexarea segmentului rezultat la cromozomul sau cromatida omoloaga. Fragmentul anexat poate avea o orientare identica cu cea a cromozomului normal, rezultand o duplicatie in tandem. Uneori inainte de anexare segmentul se roteste cu 180o rezultand o duplicatie inversa sau palindrom.
Duplicatiile pot rezulta si prin crossing-over inegal intre cromatidele unui bivalent. Ca urmare, apare un cromozom cu un segment in dublu exemplar, in timp ce celalalt va prezenta o deletie.
Inversia consta in ruperea unui fragment cromozomial urmata de sudarea acestuia in punctele de ruptura, dupa ce a suferit o rotatie de 180 . In functie de pozitia segmentului inversat, inversiile pot fi terminale sau intercalare. Acest din urma tip presupune producerea a doua rupturi. In functie de pozitia rupturilor inversiile intercalare pot fi: pericentrice, daca punctele de ruptura sunt situate pe brate cromozomiale diferite, de o parte si de alta a cetromerului, si intercalare sau paracentrice, cand cele doua puncte de ruptura sunt pe acelasi brat. In cazul inversiilor cele doua puncte de ruptura pot fi echidistante fata de centromer sau nu. In primul caz inversarea fragmentului central nu schimba pozitia centromerului ci numai a genelor pe cand in cea de-a doua situatie se schimba si pozitia centromerului. Acest tip de restructurare cromozomiala poate fi detectata relativ usor prin bandarea cromozomlor. In meioza inversiile se pot detecta deoarece in timpul sinapselor dintre cromozomii omologi segmentul inversat va forma o bucla cu segmentul omolog normal datorita inversarii succesiunii genelor. Inversiile sunt restructurari cromozomiale balansate dar duc la schimbarea succesiunii genelor pe cromozom fapt ce poate genera un efect de pozitie. Cand unul din punctele de ruptura este localizat la nivelul unei gene functionale importante efectele pot fi grave.
Citogenetica umana moderna a aparut abia dupa 1956 cand Tjio si Levan au stabilit ca numarul corect, diploid de cromozomi ai speciei umane este de 46. Tot ei au fost aceia care au descris principalele caractere morfologice ale cromozomilor umani. Omul a cunoscut deci nucleul atomului inainte de a cunoaste numarul exact de cromozomi din celulele propriului sau corp. Aceasta intarziere a fost determinata de dificultati tehnice considerabile. Singurul material disponibil era reprezentat de sectiunile histologice taiate la o grosime care sa permita pastrarea intacta a garniturii de cromozomi dintr-o celula. Cromozomii erau aglomerati iar utilizarea fotografiei era imposibila datorita numeroaselor suprapuneri. In aceste conditii singura metoda de studiu a fost reprezentata de examenul microscopic in diferite planuri focale, urmat de redarea grafica prin desene a imaginilor.
Utilizand culturi de fibroblasti derivati din plamanii unor fetusi umani avortati, Tjio si Levan au observat in mod constant un numar de 46 de cromozomi. Corectitudinea acestor observatii a fost la scurt timp confirmata de Ford si Hammerton, care pe un material provenit din biopsii testiculare au numarat 46 de cromozomi in metafazele spermatogoniilor si 23 de bivalenti in spermatocitele primare aflate in metafaza I. Aceste cercetari, cat si cele care au urmat au beneficiat de o serie de imbunatatiri tehnice dintre care cele mai importante au fost:
utilizarea culturilor de tesuturi. Celulele cultivate in vitro prezinta un numar mai mare de mitoze iar calitatea lor este superioara deoarece agentii hipotonizanti si fixatori pot actiona mai bine asupra celulelor;
pretratarea celulelor cu colchicina (sau produsul sintetic similar denumit colcemid) blocheaza in mod specific celula in metafaza ca urmare a interactiunii cu fusul de diviziune. De asemenea, colchicina contracta cromozomii permitind o mai buna individualizare a acestora;
utilizarea solutiilor hipotone determina tumefierea celulelor si permite separarea cromozomilor dispusi intr-un spatiu mai mare;
strivirea celulelor (tehnica squash) determina dispozitia cromozomilor (separati datorita tratamentului hipotonic dar aflati inca intr-o celula de forma sferica) intr-un singur plan focal si permite astfel caracterizarea morfologica a cromozomilor.
Ca urmare a imbunatatirii tehnicilor de lucru a devenit posibila nu numai numararea cromozomilor ci si studierea trasaturilor morfologice ale fiecarui cromozom in parte. In felul acesta s-a putut stabili cauza sindroamelor de trisomie 21, 13 si 18 precum si a sindroamelor Turner si Klinefelter.
Introducerea metodei autoradiografiei a permis o identificare mai precisa a cromozomilor precum si identificarea unor modificari cromozomiale minore (deletii, inversii, translocatii) ce duceau la aparitia unor sindroame clinice corespunzatoare.
In anul 1970 au aparut tehnicile de bandare cromozomiala datorita lucrarilor de pionierat ale lui Cassperson si colab. care au demonstrat ca prin colorare cu quinacrina si examinare in UV se obtine o diferentiere longitudinala a cromozomilor sub forma unei succesiuni de benzi clare si intunecate. Modelul de bandare este caracteristic pentru fiecare pereche de cromozomi omologi in parte.
CROMOZOMII UMANI STRUCTURA SI NUMAR
Cromozomii sunt structuri filamentoase localizate in nucleu. Termenul de cromozom inseamna in stricto senso corpuscul colorat si deriva din limba greaca in care soma=corp iar chroma=culoare.
Cea mai mare parte a cunostintelor privitoare la structura cromozomilor umani a fost obtinuta prin studii de microscopie optica. Coloranti speciali, absorbti specific de ADN au permis identificarea fiecarui cromozom in parte. Cromozomii sunt mai usor de observat in timpul diviziunii celulare cand ating un maxim de condensare iar genele nu mai pot fi transcrise. Lungimea cromozomilor este variabila in cursul diviziunii, considerandu-se ca sunt mai lungi in profaza si anafaza decat in metafaza
Celulele tinere (inainte de faza de replicare a ADN) detin o singura macromolecula de ADN bicatenar, formand cromozomii monocromatidici. Dupa faza S de replicare a ADN, cromozomii devin bicromatidici, posedand doua macromolecule de ADN bicatenar. Cele doua cromatide ale unui cromozom dispuse aproape paralel, sunt reunite in regiunea centromerului, regiune denumita si constrictie primara. Prin centromer se intelege o regiune distincta a cromozomului prin care acesta se ataseaza in mod reversibil de microtubulii fusului de diviziune, fapt ce-i permite sa se orienteze si sa migreze spre unul din cei doi poli ai celulei in timpul anafazei. Deoarece centromerul determina deplasarea cromozomilor el a mai fost denumit si kinetochor. Centromerul imparte cromozomul in doua: bratul lung (q) si bratul scurt (p) si are o pozitie variabila dar constanta pentru acelasi cromozom. In functie de pozitia centromerului cromozomii pot fi: metacentrici cand centromerul este localizat median iar cele doua brate sunt de lungimi aproape egale; submetacentrici cand centromerul este plasat inafara regiunii mediane iar unul din brate este mai lung decat celalalt si acrocentrici cand centromerul este plasat foarte aproape de unul din capete si cromozomul apare format din doua brate inegale dintre care unul foarte lung si altul foarte scurt.
Unii cromozomi prezinta pe langa constrictia primara si o constrictie secundara care izoleaza de restul cromozomului o portiune denumita satelit sau trabant.
Extremitatea fiecarui cromozom poarta numele de telomer. Telomerele joaca un rol esential in mentinerea stabilitatii si integritatii cromozomilor. La om, acestea sunt formate din mai multe secvente TTAGGG, repetate in tandem, care sunt mentinute de enzime denumite telomeraze. Se apreciaza ca reducerea nivelului telomerazelor asociata cu scaderea numarului de secvente TTAGGG joaca un rol important in procesele de imbatranire si moarte celulara.
Nucleul fiecarei celule somatice contine doua seturi de cromozomi, fiecare mostenit de la un parinte. Membrii unei perechi de cromozomi sunt numiti cromozomi omologi.
Materialul din care sunt formati cromozomii, ADN si proteine histonice, poarta numele de cromatina.
Gradul de condensare colorare a cromozomilor nu se realizeaza uniform. Acest lucru se observa mai ales in profaza si teleofaza. Ca urmare se apreciaza ca cromatina prezinta doua stari functionale alternative si reversibile: eucromatina si heterocromatina. Ele rezulta din faptul ca cromatina poate avea grade de condensare diferite in diferite regiuni ale cromozomilor. Gradul de condensare al fibrei de cromatina este corelat cu activitatea transcriptionala, cromatina nefiind exprimata cand se gaseste in stare de condensare maxima. Cromozomii mitotici reprezinta un caz extrem, in sensul ca ei sunt transcriptional inerti, in celula transcriptia nerealizandu-se ii timpul diviziunii celulare. Eucromatina reprezinta zonele mai slab condensate ale cromatinei, zone care se coloreaza mai putin intens cu coloranti specifici. La nivelul ei se gaseste cea mai mare parte a proteinelor nonhistonice si secventele unice de ADN; ca urmare reprezinta partea activa genetic a cromatinei interfazice. Heterocromatina reprezinta acele regiuni ale genomului care sunt ii interfaza intr-o stare de intensa condensare si nu se exprima adica este inactiva din punct de vedere transcriptional. Condensarea materialului genetic este probabil responsabila de inactivitatea sa. Trebuie sa precizam insa ca reversul acestei afirmatii nu este adevarat in sensul ca genele active se gasesc intr-adevar la nivelul eucromatinei, dar numai o mica parte din secventele eucromatice sunt transcrise. Localizarea in eucromatina este deci o conditie necesara dar insuficienta pentru exprimarea unei gene.
La om, nucleul celulelor normale contine 46 de cromozomi impartiti in perechi, existand astfel 22 de perechi de autosomi si o singura pereche de cromozomi de sex XX la femeie si XY la barbat. Celulele somatice sunt denumite diploide continand toti cei 46 de cromozomi in timp ce gametii (ovulul si spermatozoidul) sunt haploizi continand doar 23 de cromozomi.
Numarul total de cromozomi la diferite organisme variaza considerabil dar este constant pentru o anumita specie. La unele specii de feriga nucleul contine 500 de cromozomi in timp ce la o specie de crustacee se estimeaza ca in nucleu se gasesc intre 1500 si 1600 de cromozomi. La cealalta extrema se gaseste un vierme intestinal la care nucleul contine doar doi cromozomi. Unele maimute au, ca si omul, 46 de cromozomi. Primatele superioare insa, precum cimpanzeul, gorila si urangutanul au 48 de cromozomi. Dintre aceste primate cromozomii cimpanzeului seamanna cel mai bine cu cei umani, existand o diferenta de doar 1%. Se considera ca cromozomul nr. 2 de la om ar fi rezultatul unei fuziuni a doi cromozomi de la cimpanzeu, celelalte diferente intre complementul cromozomial de la cele doua specii fiind datorate numeroaselor inversii peri- si paracentrice. Aceste observatii au fost exploatate de specialistii in biologie moleculara pentru cartarea si clonarea genelor umane.
Cromozomii X si Y sunt cunoscuti sub denumirea de cromozomi ai sexului datorita rolului lor crucial in determinarea sexelor. La om precum si la majoritatea mamiferelor atat femela cat si masculul au doi cromozomi de sex- XX la femela si XY la mascul. Cromozomul Y este mult mai mic decat X si poarta doar cateva gene de importanta functionala, cea mai importanta fiind SRY sau factorul de determinare testiculara.
La femeie fiecare ovul poarta cate un cromozom X, pe cand la barbat spermatozoidul poate purta ori X ori Y. Sansa ca un spermatozoid ce poarta cromozomul X sau Y sa fecundeze un ovul este aproximativ egala, asa cum si numarul de produsi de conceptie de sex masculin si feminin este aproximativ egal. In realitate, se nasc mai multi copii de sex masculin ca si cei de sex feminin desi in timpul copilariei si maturitatii sex ratio-ul ajunge 1:1.
METODE DE ANALIZA CROMOZOMIALA
Cu unele imbunatatiri, metodele folosite de Tjio si Levan sunt folosite si astazi in aproape toate laboratoarele de citogenetica in scopul realizarii unei analize a constitutiei cromozomiale a unui individ. In acest sens se realizeaza cariotipul, adica aranjarea sistematica standardizata a cromozomilor unei celule, fotografiati la microscop in timpul metafazei mitozei.
PREPARAREA CROMOZOMILOR
Pentru a studia cromozomii umani pot fi folosite practic orice tesuturi cu celule vii, nucleate care se pot divide. Cel mai frecvent sunt folosite limfocitele din sangele periferic desi probe pentru analiza cromozomiala pot fi preparate relativ usor folosind piele, maduva hematogena, vili corionici sau amniocite( celule provenite din lichidul amniotic).
In cazul in care pentru obtinerea preparatelor cromozomiale se foloseste sangele periferic (venos), limfocitele sunt mai intai separate si trecute apoi pe un mediu de cultura la care se adauga fitohemaglutinina care stimuleaza limfocitele T sa se divida. Celulele sunt tinute in conditii sterile la 37oC aproximativ 3 zile, timp in care ele se vor divide. Inainte de realizarea preparatelor se adauga colchicina. In final celulele sunt separate din cultura si tratate cu solutie hipotona care produce umflarea celulelor si dispersia cromozomilor. Preparatele sunt apoi fixate pe o lama de microscop iar dupa colorare vor putea servi analizei citogenetice.
Tehnicile conventionale asigura o colorare uniforma, nebandata a cromozomilor. Ele sunt la ora actuala mai putin folosite datorita apaitiei posibilitatii de bandare cromozomiala. Bazele chimice ale reactiilor de colorare sunt inca insuficient cunoscute.
Prin tehnicile de bandare de-a lungul cromozomilor apare o succesiune de benzi transversale. Aceasta alternanta de benzi clare cu benzi intunecate este identica la cromozomii omologi si diferita pentru cromozomii apartinand diferitelor perechi de cromozomi omologi.
Bandarea Giemsa
Bandarea Giemsa a devenit astazi una dintre cele mai cunoscute si utilizate tehnici de bandare pentru cromozomii mamiferelor. Cele mai uzuale metode pentru a obtine aceasta colorare se bazeaza pe tratarea lamelor cu preparate cromozomiale cu o proteaza (de regula tripsina) sau pe incubarea lamelor in solutie citrat salina fierbinte, desi exista o mare varietate de alte metode. Amprenta cromozomului bandat se apreciaza ca reflecta atat compozitia sa structurala, cat si cea functionala. Benzile inchise la culoare, corelate cu cromomerele din pachiten, isi replica ADN-ul in faza S tarzie, si contin portiuni bogate in T-A. Se pare ca ele contin putine gene active si pot diferi de benzile deschise la culoare prin proteinele cromozomiale. Extractia diferentiata a proteinelor in pretratamentul de fixare si bandare este importanta in mecanismul de obtinere a benzilor G.
Bandarea Q
Quinacrina este un colorant acridinic care se leaga de ADN fie de ionii externi, fie prin intercalare. Bandarea in acest sistem pare a fi puternic influentata de variatia in compozitia proteica a cromozomilor. Bandarea cu quinacrina duce la aparitia benzilor Q care sunt asemanatoare ca dimensiuni si dispozitie benzilor G. Acest tip de bandare permite insa o mai buna evidentiere si analiza a unor portiuni cromozomiale de mare importanta precum regiunile satelitare ale cromozomilor acrocentrici si regiunile centromerice ale cromozomilor 1, 9 si 16 . Polimorfismul acestor regiuni, ca si variatiile cromozomului Y, pot fi mai usor observate cu un astfel de sistem de bandare.
Bandarea cu quinacrina este relativ simpla, rapida si nu necesita pretratamente speciale inainte de colorare. Metoda da rezultate bune chiar si cu lame mai vechi sau chiar in cazul unor preparate cromozomiale mai putin reusite.
Bandarea de reversie
Benzile care apar ca deschise la culoare prin bandarea Giemsa, vor aparea ca avand o coloratie intensa prin bandare de reversie. Benzile R pot fi obtinute prin incubare in solutie salina fierbinte inainte de colorare Giemsa, sau prin colorare cu acridin oranj. O alta metoda implica colorarea cromozomilor cu cromomicina A (un colorant fluorescent), urmata de o a doua colorare cu verde de metil care se leaga de perechile de baze azotate complementare celor de care se leaga cromomicina A. Ca urmare, vor ramane fluorescente doar regiunile in care perechile de baze sunt predominant de un singur tip.Studiul in microscopia electronica de tip scanning a cromozomilor umani bandati G releva in cromatidele cromozomilor regiuni proeminente care alterneaza cu santuri circulare.Santurile circulare corespund benzilor G pozitive iar regiunile proeminente corespund benzilor R pozitive observate in microscopia optica.
Bandarea C (heterocromatina centromerica)
Acest tip de bandare se obtine dupa tratamente de denaturaresi renaturare ale cromozomilor urmate de colorare cu solutie Giemsa. Se obtine astfel o coloratie intensa a heterocromatinei constitutive din regiunile pericentromerice care sunt caracteristice in ce priveste intinderea pentru fiecare cromozom in parte. Cromozomii 1,9 si 16 contin cate un segment mare de heterocromatina extins in bratul q. romosomul romosomul Y prezinta un segment heterocromatic in jumatatea distala a bratului q ce corespunde segmentului q stralucitor si o banda ingusta la nivelui centromerului.Satelitii au o coloratie mai fina decat regiunile centromerice.Bandarea C permite evidentierea variatiilor individuale sau familiale ale constrictiilor secundare si ale heterocromatinei pericentromerice precum si detectarea centromerilor in cazurile de remanieri cromozomiale. Aparitia ocazionala de benzi C cu localizare terminala sau intercalara are la origine dislocarea heterocromatinei din regiunea centromerica prin inversii sau translocatii.
Bandarea T
Benzile T (T=telomere) se obtin in urma unei denaturari termice mai pronuntate necesara pentru suprimarea benzilor R de pe bratele cromozomilor cu exceptia telomerelor. Aceste benzi reprezinta fractiunea cea mai rezistenta la tratamentul termic al benzilor R. Dupa colorare cu acridin orange cromozomii observati in lumina UV prezinta o coloratie slaba portocalie iar telomerele au o fluorescenta verde a carei intensitate variabila permite diferentierea cromozomilor intre ei. Acest tip de mercaj este util intrucat permite depistarea unor modificari minore ce vizeaza extremitatile cromozomiale.
Bandarea de rezolutie inalta
Bandarea G permite in mod normal identificarea a circa 400 de benzi per set haploid de cromozomi umani. Fiecare dintre aceste benzi corespunde la circa 8000 kb de ADN. Bandarea de rezolutie inalta a cromozomilor aflati in profaza sau prometafaza (cand au un nivel de condensare mai scazut) permite evidentierea a circa 800 de benzi per set haploid de cromozomi. Aceasta tehnica necesita inhibarea diviziunii celulare cu un antagonist al acidului folic precum metotrexatul. Acidul folic este adaugat mediului de cultura pentru a permite intrarea celulelor in mitoza.Ulterior este adaugata colchicina la un anumit interval de timp, si anume atunci cand cea mai mare parte din celule vor fi in prometafaza.
ANALIZA CROMOZOMIALA
In vederea realizarii cariotipului cromozomii trebuie mai intai identificati cu precizie si aranjati in perechi. Caracterizarea morfologica a cromosmilor metafazici are la baza o serie de criterii stabilite de ISCN-International System for Human Cytogenetic Nomenclature.
1.Lungimea relativa a unui cromozom este reprezentata de raportul dintre
lungimea cromozomului( reprezentata de suma lungimii bratelor p+q) si lungimea totala a unui set haploid de cromozomi(22 de autosomi +X).
Lungimea absoluta a cromozomilor nu are o importanta prea mare pentru caracterizarea acestora deoarece variaza in functie de stadiul metafazei, cromozomii fiind mai lungi in metafaza precoce (prometafaza) decat in metafaza tardiva. Agentii statmochinetici utilizati in mod curent in citogenetica pentru blocarea mitozelor in metafaza (colchicina, colcemid, vinblastina) accentueaza aceasta scurtare.
2. Indicele centromeric este definit prin raportul dintre bratul scurt (p) si lungimea totala a cromozomului.
IC=p/(p+q)
3. Raportul bratelor este reprezentat de dimensiunea bratului lung (q) raportata la dimensiunea bratului scurt (p).
B=q/p
La cromozomii metacentrici raportul este de 1.0-1.7, la cei submetacentrici 1.7-3.0, la subtelocentrici 3.0-7.0 iar la cei acrocentrici 7.0-4
4.Existenta satelitilor. Satelitii sunt prezenti la cromozomii acrocentrici si sunt reprezentati de mase mici de cromatina. Ei se gasesc la extremitatea terminala a bratelor scurte a cromozomilor de care sunt separati prin constrictii secundare.
5.Constrictii secundare. Termenul de constrictie primara poate fi considerat sinonim cu cel de centromer. Unii cromozomi prezinta insa constrictii si in alte regiuni ale bratelor, denumite constrictii secundare.
6.Replicarea asincrona a cromozomilor. Evidentierea replicarii asincrone a cromozomilor umani in cadrul perioadei S a ciclului celular a fost posibila in urma administrarii de H3 timidina si urmarirea incorporarii acesteia in cromozomi in functie de momentul replicarii. Metoda a permis deferentierea cromozomilor din grupele B,D si E. De asemenea s-a pus in evidenta replicarea tardiva a unuia din cei doi cromozomi X la femei.
7.Modelul de benzi cromozomiale se refera la obtinerea unor coloratii diferentiale ale cromozomilor.
Pe baza criteriilor mentionate, cromozomii umani au fost repartizati in 7 grupe morfologice.
Grupa A (1-3). Cuprinde cromozomi mari (9,08-7,06F) dintre care 1 si 3 sunt metacentrici iar cei din perechea 2 sunt submetacentrici. Cromozomii din perechea 1 prezinta o constrictie secundara in regiunea proximala a bratului q.
Grupa B (4-5). este alcatuita din cromozomi lungi submetacentrici si au dimensiuni situate intre 6.55-6,13F
Grupa C (6-12) si X cuprinde cromozomi de marime mijlocie (5,84-4,46F). Perechile 6,7,8,11 au centromerul situat in regiunea mediana (sunt metacentrici) iar perechile 9,10 si 12 au centromerul situat submedian deci sunt submetacentrici. Cromozomii din perechea 9 prezinta o constrictie secundara in regiunea proximala a bratului q. Cromozomii X sunt metacentrici si au dimensiuni similare cu a cromozomilor din perechile 7 si 8.
Grupa D (13-15) cuprinde cromozomi de marime mijlocie (3,64-3,36F) acrocentrici. Toti cromozomii acestei grupe prezinta sateliti.
Grupa E (16-18) cuprinde cromozomi relativ mici (3,23-2,76F) cu centromerul situat median la cei din perechea 16 si submedian la perechile 17 si 18.Cromozomii din perechea 16 au o constrictie secundara in regiunea proximala a bratului q.
Grupa F (19-20) cuprinde doua perechi de cromozomi mici (2,52-2,33F), metacentrici.
Grupa G (21-22) cuprinde cei mai mici cromozomi ai cariotipului uman (1,83-1,68F), sunt acrocentrici si prezinta sateliti.
Cromozomul Y are lungimea apropiata de cea a cromozomilor din grupa G (1.96F), este de tip acrocentric dar spre deosebire de ceilalti cromozomi acrocentrici din cariotip nu au sateliti.
HIBRIDIZARE FLUORESCENTA IN SITU (FISH)
Metodele noi de diagnostic combina tehnicile citogenetice conventionale cu cele de inginerie genetica moleculara. Tehnica FISH se bazeaza pe capacitatea unei portiuni de ADN monocatenar de a se lega de o secventa complementara care poate fi localizata oriunde pe o placa metafazica. Spre deosebire de alte metode folosite in analiza cromozomilor tehnica FISH poate sa fie aplicata si celulelor aflate in interfaza.
In aceasta tehnica proba de ADN este conjugata cu nucleotide modificate care dupa hibridizare cu ADN provenit de la pacient permite regiunii unde a avut loc hibridizarea sa devina vizibila in lumina UV. Aceasta tehnica este larg folosita astazi in scopul diagnosticarii clinice si datorita faptului ca rezultatele se obtin foarte rapid.
POLIMORFISMUL CROMOZOMILOR UMANI
Studiile au aratat ca anumite regiuni ale cromozomilor prezinta variatii ale dimensiunii sau ale afinitatii fata de coloranti. Intrucat acest polimorfism nu are o expresie fenotipica deci nu exercita nici o influenta asupra fenotipului, diferentele structurale mentionate sunt considerate variante normale.
Variabilitatea cromozomiala implica regiunile constrictiilor secundare, portiunea distala a cromozomului Y si satelitii. Benzile C si Q pot prezenta variatii ale lungimii in cadrul diferitelor grupe de cromozomi, fiind caracteristice pentru fiecare individ in parte.Benzile mai mari rezulta din repetarea in tandem a unor unitati mai mici, identice. Cromozomii 1, 9, 16 au benzi care variaza atat in lungime cat si in localizarea lor in regiunea centromerica si a constrictiilor secundare. In majoritatea cazurilor banda este extinsa in bratul lung si foarte rar in bratul scurt ca rezultat al unor inversii pericentrice. Variatia in lungime a benzilor C in acesti cromozomi se datoreaza in primul rand variatiei in lungime a constrictiilor secundare.
Cromozomul Y are o banda heterocromatica foarte ingusta la nivelul centromerului, pentru care diferentele de lungime de la un cromozom la altul sunt dificil de raportat. Segmentul distal al bratului q poate insa varia considerabil in lungime.
Au fost semnalate urmatoarele tipuri de variante cromozomiale:
Constrictie secundara alungita la cromozomii 1, 9, 16. Se considera constrictia secundara ca fiind alungita atunci cand are o lungime cel putin dubla fata de lungimea constrictiei secundare a cromozomului omolog.
Bratul p alungit la cromozomii acrocentrici. Bratul p se considera alungit atunci cand este egal sau mai lung decat bratul scurt al unui cromozom din perechea 18.
Sateliti proeminenti. Sunt definiti astfel cand sunt mai mari decat bratul scurt de la acelasi cromozom.
Sateliti dubli- se remarca prin colorare doi sateliti in tandem.
Cromozom y mare sau mic. Cromozomul Y este considerat mare atunci cand Y/F>1,00 si mic atunci cand raportul este de sub 0,7.
Prezenta satelitilor pe bratul scurt al cromozomului 17 (17ps). Aceasta varianta se intalneste foarte rar, semnalandu-se in cateva cazuri printre pacientii internati in clinicile de psihiatrie.
Prezenta satelitilor pe bratul q al cromozomului y. Este o varianta foarte rar intalnita fiind semnalata fie la pacienti normali fie la pacienti cu sindromul Klinefelter.Acest caz particular se pare ca rezulta in urma translocatiei satelitilor unui cromozom din grupele D sau G pe bratul q al cromozomului Y.
ANOMALII (ABERATII CROMOZOMIALE)
O mare parte a maladiilor conditionate genetic este datorata aberatiilor (mutatiilor) cromozomiale numerice sau structurale.
Aberatii numerice
Aberatiile numerice pot fi impartite in doua mari categorii si anume:
3. aneuploidia - datorata pierderii sau dobandirii unuor cromozomi suplimentari;
4. poliploidia datorata aditiei unuia sau a doua seturi haploide complete
1. Aneuoloidia
Cele mai frecvent intalnite aneuploidii sunt trisomiile datorate aparitiei unui cromozom suplimentar, tetrasomiile datorate existentei in celule a doi cromozomi suplimentari pentru o anumita pereche de cromozomi omologi si monosomiile rezultate ca urmare a pierderii unui cromozom.
Sindromul Down este datorat unui cromozom suplimentar din perechea 21 si ca urmare este cunoscut si sub numele de trisomie 21. Exista si alte trisomii ale autosomilor compatibile cu supravietuirea precum sindromul Patau (trisomia 13) si sindromul Edwards (trisomia 18). Cele mai multe trisomii ale autosomilor duc insa la pierderea timpurie a sarcinii, trisomia 16 fiind relativ frecvent intalnita in cazul avorturilor spontane in primul trimestru al sarcinii. Prezenta in exces a unui heterosom (X sau Y) are efecte fenotipice mai putin grave.
Originea aneuploidiei
Principalele cauze ale aneuploidiei sunt: non-disjunctia, pierderea de cromozomi datorita retardarii anafazice, endoreduplicarea selectiva si mitozele multipolare.
Non-disjunctia este procesul prin care cromozomii omologi (sau cromatidele aceluiasi cromozom) nu se separa pentru a se distribui in mod egal celulelor fiice, ci migreaza impreuna in una din aceste celule. Acest proces poate avea loc atat in timpul gametogenezei cat si in primele stadii ale clivarii zigotului.
Procesul de non-disjunctie poate aparea in meioza I, meioza II sau (mai rar) in ambele diviziuni meiotice. Vorbim despre non-disjunctie primara cand celulele germinale sunt normale si non-disjunctie secundara ce opereaza pe celule germinale anormale (de ex. 47, xxx).
Consecintele non-disjunctiei sunt diferite la barbati si la femei, deosebire determinata de particularitatile gametogenezei la cele doua sexe. Astfel la barbat dintr-un spermatocit primar rezulta 4 spermatozoizi functionali; la femeie dintr-un ovocit primar rezulta un singur ovul functional, fiecare diviziune meiotica fiind insotita de eliminarea unui globul polar. Din acest motiv la barbati non-disjunctia in decursul meiozei I va determina aparitia unor spermatozoizi disomici si nulisomici in timp ce non-disjunctia in meioza II va produce si gameti normali alaturi de gameti disomici si nulisomici.. La femeie insa non-disjunctia survenita fie in decursul primei, fie a celei de-a doua meioze nu poate produce decat ovule anormale: disomice sau nulisomice. Posibilitatea ca un zigot anormal sa fie consecinta unei anomalii survenite in timpul ovogenezei este deci mai mare decat cea a unei anomalii similare aparute in decursul spermatogenezei. Studiile au aratat ca cea mai mare parte a copiilor cu trisomii autosomaleau mostenit cromozomul suplimentar de la mama (Tab. )
Tabelul nr. Originea cromozomuli suplimentar in cazul celor mai frecvente trisomii
|
Tipul aberatiei cromozomiale |
Origine paterna (%) |
Origine materna (%) |
|
Trisomia 13 |
15 |
85 |
|
Trisomia 18 |
10 |
90 |
|
Trisomia 21 |
5 |
95 |
|
45,XXX |
5 |
95 |
|
47,XXY |
45 |
55 |
|
47,XYY |
100 |
0 |
|
|
|
|
Cu o frecventa redusa, non-disjunctia poate sa apara si in primele diviziuni mitotice ale zigotului. Vor rezulta doua sau mai multe linii celulare diferite, fenomen denumit mozaicism. Constitutia cromozomiala a organismului care rezulta depinde de momentul in care are loc non-disjunctia. Astfel, daca non-disjunctia are loc in timpul primei diviziuni de clivare a unui zigot normal, va rezulta un produs de conceptie care va prezenta doua linii celulare anormale, una monosomica si una trisomica pentru cromozomul in cauza. Daca non-disjunctia apare in a doua diviziune a zigotului, va rezulta un blastocit mixoploid alcatuit din trei linii celulare: una normala disomica, una trisomica si una monosomica. In cursul vietii indivizilor se poate produce o selectie care va duce la eliminarea celulelor dezavantajate (in special a celor monosomice). Intrucat aceasta selectie opereaza mai eficient in tesuturile cu diviziune rapida, proportia celulelor anormale nu va fi identica in toate tesuturile unei anumite persoane.
Cauzele non-disjunctiei
Cauzele non-disjunctiei nu sunt inca suficient de bine cunoscute. Evenimentele non-disjunctionale pot aparea in decursul ambelor diviziuni meiotice ale spermatogenezei si ovogenezei, dar survin cu o frecventa mai mare in ovogeneza. In acest sens pledeaza faptul ca varsta medie a mamelor care au dat nastere unor copii ci trisomii 21, 13 sau 18, este mai ridicata decat a mamelor care au dat nastere unor copii normali (tab. de la S.Down). Acest fenomen ar putea fi explicat prin faptul ca spermatogeneza are loc continuu, in tot cursul vietii, in timp ce la femeie prima diviziune meiotica are loc inainte de nastere, ovocitele ramanand in ovar in stare de repaus pana in momentul ovulatiei care are loc intre 15-50 de ani. In tot acest interval de timp ovocitele sunt supuse unui proces de imbatrinire si unor influente externe (iradieri, infectii, medicamente) care pot reduce eficienta meiozei. Studiile au aratat, de asemenea, ca absenta recombinarii in profaza meiozei I ar predispune la non-disjunctie. Aceasta observatie nu este surprinzatoare deoarece chiasmata sunt responsabile de mentinerea conjugata a celor doi cromozomi omologi pana in diakineza cand se vor separa. La femeie recombinarea se produce inainte de nastere in timp ce non-disjuctia se produce cu 15-50 de ani mai tarziu. Aceasta observatie sugereaza faptul ca non-disjunctia ar putea fi cauzata de cel putin doi factori, si anume: absenta recombinarii intre cromozomii omologi in perioada fetala si aparitia, mult mai tarziu, unor anomalii in formarea fusului de diviziune.
O alta explicatie a aocierii intre varsta avansata a mamei si riscul nasterii unui copil afectat de o trisomie autosomala, ar consta in aceea ca supravietuirea embrionilor trisomici ar putea fi rezultatul unei diminuari a raspunsului imun al mamei, odata cu varsta. Se apreciaza, de asemenea, ca non-disjunctia ar fi favorizata si de cresterea intervalului de timp dintre ovulatie si fertilizare.
La unele specii precum cele ale genului Drosophila, s-a constatat ca non-disjunctia este controlata genetic. Aceasta observatie ar putea furniza o explicatie pentru familiile care par a fi predispuse la non-disjunctii repetate.
Procesul de retardare anafazica reprezinta o alta cauza a pierderii de cromozomi. In decursul anafazei, unul sau cativa cromozomi nu se deplaseaza spre polii celulei astfel incat dupa ce membrana celulara se reface in telofaza ei sunt exclusi din nucleu, pierzandu-se in citoplasma.
Endoreduplicarea selectiva a fost definita ca o replicare dubla a unei parti din genom (un cromozom intreg sau o parte de cromozom).
Mitozele multipolare sunt o cauza a anuploidiilor In special in celulele canceroase. Ele sunt consecinta formarii unui fus multipolar datorat unor diviziuni succesive ale centrosomului si duc la repartizari inegale ale cromozomilor in celulele fiice.
Un procent relativ mic din numarul de aneuploizi ar putea fi atribuit si altor factori precum:
tulburarilor autoimune parentale (anticorpi anttiroidieni la mama);
actiunea mutagena a iradierilor medicale;
infectiile virale materne.
2. Poliploidia
Organismele poliploide contin in celule un multiplu al numarului haploid de cromozomi. La om, formele triploide contin 69 de cromozomi in celule si au fost intalnite in special in cazul analizelor citogenetice a produsilor de conceptie avortati. Literatura de specialitate descrie doar cateva cazuri de nou-nascuti triploizi, dar acestia au murit la scurt timp dupa nastere. Zigotii triploizi pot lua nastere fie prin fecundarea unui ovul de catre doi spermatozoizi (dispermie), fie, prin fuziunea ovulului cu un globul polar (diginie). S-a constatat ca in cazul triploizilor rezultati prin dispermie placenta este marita, hidatidiforma in ce in cazul celor rezultati in urma dispermiei placenta este mai mica decat in situatii normale.
Tetraploizii au la om 92 de cromozomi si apar cu o frecventa foarte mica, tetraploidia fiind cosiderata incompatibila cu viata. Astfel de constitutii genetice au fost identificate in urma realizarii analizei cromozomiale a unor produsi de conceptie avortati spontan. Se apreciaza ca aceste forme apar, cel mai probabil, ca urmare a unei supresii a primei diviziuni de clivare a unui zigot diploid, dupa separarea cromozomilor dar inaite de diviziunea citoplasmei.
Poliploidizarea poate aparea si in decursul vietii adulte datorita unor accidente mitotice (endomitoza sau endoreduplicarea). Celule poliploide au fost evidentiate in proportie foarte redusa in maduva osoasa si in culturile de limfocite, fiind insa foarte frecvente in celulele tumorale. In cazul endomitozei cromozomii se comporta normal pana in profaza tarzie cand membrana nucleara nu se dezintegreaza iar fusul de diviziune nu se formeaza. Endoreduplicarea se caracterizeaza prin doua duplicari succesive a cromozomilor in interfaza astfel ca in profaza si metafaza se observa un numar dublu de cromozomi. Procesul de separare a cromozomilor se desfasoara in mod normal dar el opereaza pe o garnitura cromozomiala dubla si va duce in final la aparitia a doua celule tetraploide.
Aberatii structurale
Aberatiile structurale ale cromozomilor rezulta ca urmare a aparitiei unor rupturi urmate fie de pierderea segmentului lipsit de centromer, fie de realipirea fragmentului rupt intr-o configuratie diferita.
Aberatiile cromozomiale pot fi balansate sau nebalansate. In cazul celor balansate complementul cromozomial ramane complet deoarece nu a avut loc o pierdere sau un castig de material genetic sau nebalansate. Ca urmare acest tip de aberatii cromozomiale nu are in general urmari mai putin rarele situatii in care unul din punctele de ruptura este localizat la nivelul unei gene functionale importante. Purtatorii unor aberatii cromozomiale balansate prezinta un risc de a avea copii cu un complement cromozomial anormal. In cazul aberatiilor cromozomiale nebalansate, se produce o modificare a cantitatii de material genetic iar urmarile asupra organismului sunt cel mai adesea severe.
Rupturile
Constau in intreruperi ale continuitatii cromatidelor sau cromozomilor insotite de o deplasare laterala a fragmentelor distale. Ele pot avea trei consecinte:
reunirea fragmentelor cu refacerea structurii initiale a cromozomilor (restitutie);
reunirea cu modificare rezultand diferite tipuri de aberatii cromozomiale;
lipsa reunirii cu pierderea fragmentului rupt.
Agentii care provoaca rupturi cromozomiale (agenti clastogeni) sunt destul de variati. Ei pot fii clasificati in factori externi (radiatii, substante chimice, virusuri) si interni (afectiuni ereditare, imbatranirea). Expunerile la doze mari de radiatii pot fi accidentale sau datorate iradierilor profesionale sau medicale. In grupa agentilor chimici intra substante extrem de numeroase utilizate pe scara larga (chimioterapicele antitumorale, unele antibiotice, pesticide, detergenti si aditivi din industria cosmetica sau alimentara). Consecintele actiunii virusurilor infectioase (virusurile hepatitei A si B, rubeolei) sau oncogene (polyoma, SV-40) sunt extrem de grave iar tabloul modificarilor cromozomiale este complex. In unele afectiuni ereditare precum Sindromul Bloom sau anemia Fanconi exista in mod spontan un nivel crescut de rupturi cromozomiale. Persoanele care sufera de aceste boli prezinta o incidenta crescuta a anumitor tipuri de cancer.
Analizand rupturile aparute spontan in limfocitele umane s-a inregistrat un mare numar de puncte de ruptura care au fost catalogate in functie de localizarea lor in benzile negative sau pozitive. Din studiile amintite mai sus se desprind urmatoarele aspecte:
rupturile spontane ca si cele induse sunt localizate de preferinta in benzile G sau Q negative;
rupturile dintr-un tip de banda tind sa se reuneasca cu cele apartnand aceluiasi tip de banda (negativ-negativ, pozitiv-pozitiv), combinatiile heterologe fiind rar intalnite;
frecventa rupturilor este mai mica in regiunile centromerice si telomerice;
distributia rupturilor nu este proportionala cu lungimea cromozomilor. Cel mai mare numar de rupturi pe unitatea de lungime o prezinta in ordine cromozomii 21, Y, 18, 9 si 13, iar cel mai mic numar de rupturi s-a observat la cromozomii 16, 6, 2, 3, 19 si 20.
Deletiile
Deletiile (lipsurile) constau in piederea unui fragment de cromatida sau de cromozom. Ele pot fi terminale sau interstitiale si sunt cauzate de rupturi care apar la nivelul cromozomului. Daca ruptura a avut loc inainte de faza S a ciclului celular va genera o deletie cromozomiala, iar dupa aceasta faza o deletie cromatidica. In urma unor deletii izocromatidice suprafetele de ruptura de la capetele cromatidelor pot fuziona, iar in faza urmatoare a diviziunii, cand cei doi centromeri rezultati prin clivare migreaza spre cei doi poli ai fusului de diviziune, cromatidele raman unite prin zona de fuzionare realizand o punte anafazica. In anafaza, fragmentele telomerice acentrice nu se pot deplasa spre unul dintre polii celulei datorita faptului ca sunt lipsite de centromer. Din aceasta cauza se pierd si rezulta o lipsa terminala simpla. Daca se produc rupturi simultane la ambele capete ale unui cromozom, zonele acentrice se pierd, iar fragmentul cu centromeri formeaza un cromozom circular (inelar) prin unirea capetelor lipsite de telomere care devin instabile si au tendinta de a se uni.
Deletiile sunt datorate, cel mai frecvent, formarii unei bucle de incalecare, la nivelul careia se produce o ruptura dubla rezultand patru capete. Va rezulta un fragment acentric (care la nivelul metafazei formeaza micronucleii) si un cromozom cu deletie. Contrar asteptarilor deletiile intercalare sunt mai frecvente decat cele terminale, deoarece nu implica formarea de noi telomere pentru stabilizarea deletiilor.
Deletia consta in pierderea un fragment de cromozom si prin urmare va duce la aparitia unei monosomii pentru fragmentul pierdut. In general o deletie mare (mai mare de 2%dintr-un set haploid de cromozomi) este considerata incompatibila cu supravietuirea.
Unele deletii, precum cele care duc la aparitia sindroamelor Wolf-Hirschhorn sau cri-du chat pot fi observate la microscop in preparate obtinute din placi metafazice. In cazul in care deletia implica un fragment de dimensiuni foarte mici identificarea se poate face numai prin realizarea unei analize citogenetice in prometafaza si studiilor de hibridizare in situ. Asa este cazul sindroamelor Angelman si Prader-Wili.
Duplicatiile
Duplicatia consta in dublarea unui fragment cromozomial sau
cromatidic.
Fig. nr.
Aparitia unei duplicatii este consecinta unei rupturi urmate de anexarea segmentului rezultat la cromozomul sau cromatida omoloaga. Fragmentul anexat poate avea o orientare identica cu cea a cromozomului normal, rezultand o duplicatie in tandem. Uneori inainte de anexare segmentul se roteste cu 180o rezultand o duplicatie inversa sau palindrom.
Duplicatiile pot rezulta si prin crossing-over inegal intre cromatidele unui bivalent. Ca urmare, apare un cromozom cu un segment in dublu exemplar, in timp ce celalalt va prezenta o deletie.
Inversia consta in ruperea unui fragment cromozomial urmata de sudarea acestuia in punctele de ruptura, dupa ce a suferit o rotatie de 180 . In functie de pozitia segmentului inversat, inversiile pot fi terminale sau intercalare. Acest din urma tip presupune producerea a doua rupturi. In functie de pozitia rupturilor inversiile intercalare pot fi: pericentrice, daca punctele de ruptura sunt situate pe brate cromozomiale diferite, de o parte si de alta a cetromerului, si intercalare sau paracentrice, cand cele doua puncte de ruptura sunt pe acelasi brat. In cazul inversiilor cele doua puncte de ruptura pot fi echidistante fata de centromer sau nu. In primul caz inversarea fragmentului central nu schimba pozitia centromerului ci numai a genelor pe cand in cea de-a doua situatie se schimba si pozitia centromerului. Acest tip de restructurare cromozomiala poate fi detectata relativ usor prin bandarea cromozomlor. In meioza inversiile se pot detecta deoarece in timpul sinapselor dintre cromozomii omologi segmentul inversat va forma o bucla cu segmentul omolog normal datorita inversarii succesiunii genelor. Inversiile sunt restructurari cromozomiale balansate dar duc la schimbarea succesiunii genelor pe cromozom fapt ce poate genera un efect de pozitie. Cand unul din punctele de ruptura este localizat la nivelul unei gene functionale importante efectele pot fi grave.
Un individ care prezinta o inversie pericentrica poate produce gameti neechilibrati genetic daca crossing-overul are loc pe portiunea segmentului inversat. Ca urmare vor rezulta doi cromozomi omologi recombinati dupa cum urmeaza: unul cu o duplicatie in portiunea segmentului distal neinversat si o deletie la celalalt capat al cromozomului iar celalalt cu o deletie la nivelul segmentului neinversat si o duplicatiela celalalt capat al cromozomului (FIG. p.50). Cu cat dimensiunea segmentului implicat in duplicatie sau deletie este mai mare cu atat sansa unui avort spontan este mai mare. Prin urmare, cu cat segmentul inversat este mai mic cu atat masura in care gametii vor fi neechilibrati genetic va fi mai mare iar sansa unui avort spontan (in cazul in care acesti gameti au contrivuit la fecundare) creste.
Indivizii care prezinta inversii paracentrice vor produce, de asemenea, gameti neechilibrati genetic daca apare un crossing-over la nivelul segmentului inversat. In acest caz va rezulta un cromozom acentric si unul dicentric (cu doi centromeri) (FIG-p50). Cromozomul acentric se va pierde (sub forma de micronucleu) la urmatoarea diviziune mitotica iar cel dicentric va avea un grad crescut de instabilitate astfel incat embrionii vor avea sanse minime de supravietuire.
Izocromozomii sunt cromozomi metacentrici cu bratele identice structural si functional, rezultati in urma unor clivari transversale a centromerului. Cel mai frecvent intalnit izocromozom este cel alcatuit din doua brate lungi ale cromomosomului X. El este raspunzator de cca. 20% din cazurile de sindrom Turner.
Fragmentele acentrice rezulta in urma unor deletii intercalare sau terminale si sunt detectabile in metafaza. Apartenenta acestora se poate stabili prin aplicarea tehnicilor de bandare. La sfarsitul mitozei fragmentele acentrice formeaza unul sau mai multi micronuclei.
Un cromozom inelar rezulta ca urmare a fuziuonarii suprafetelor de ruptura ale cromatidelor unui cromozom in urma unor deletii terminale. Ei sunt insotiti cel mai adesea de unul sau mai multe fragmente acentrice. Cromozomii inelari sunt cel mai adesea incapabili de a trece printr-o diviziune mitotica, astfel incat ei pot fi intalniti numai la o parte din celule. Celelalte celule ale individului sunt monosomice datorita absentei cromozomului inelar.
Cromozomi minusculi dubli (dms)
Acesti cromozomi au aspectul unor sfere mici de cromatina apropiate intre ele si au fost observati prima data in celule provenite din tumori umane. Forma si marimea lor prezinta o mare heterogenitate in diferitele populatii celulare tumorale. Observandu-se comportarea acestora, s-a constatat ca in profaza si in prometafaza cele doua sfere de cromatina sunt mai apropiate, in metafaza si anafaza distanta dintre ele se mareste, in telofaza se despart iar in interfaza apar ca niste micronuclei. Faptul ca dms se gasesc numai in tumori, se pot intretine si chiar inmulti, a condus la afirmatia ca ele au rol in evolutia tulpinilor maligne pentru a provoca prabusirea mijloacelor de aparare ale gazdei. Aceasta afirmatie are la baza observatia ca dms se gasesc intr-un numar mare in celulele tumorii primare iar apoi se elimina treptat in timpul cultivarii in vitro. Reimplantarea in vivo determina restabilirea populatiei de dms, iar celulele cu dms prezinta un avantaj selectiv in vivo si dezavantaj in vitro. Prin tehnicile de bandare s-a constatat ca dms nu prezinta centromeri, fapt care face greu de explicat modul in care acestia pot ramane timp indelungat in populatiile celulare maligne stiut fiind faptul ca formatiunile acentrice nu se include in nucleu la sfarsitul mitozei.
Regiuni omogen colorate (HSR)
HSR constituie un tip particular de aberatii cromozomiale ce se gasesc predominant in celulele canceroase. Prin tehnicile de bandare G acestea apar ca regiuni extinse cu intensitate de colorare scazuta sau medie. Se apreciaza ca acestea reprezinta situsuri de amplificare a unor gene specifice (oncogene). Identificarea precisa a acestor regiuni este de o deosebita importanta in vederea identificarii genelor amplificate si a functiilor lor.
Pulverizatiile cromozomiale constau intr-o fragmentare totala a cromozomilor unei celule si duc la aparitia unor granule fine, punctiforme de cromatina. Au fost identificate in celulele expuse la agenti chimici si virali.
Translocatiile constau in transferul unui segment dintr-un cromozom la un alt cromozom nereciproc. Atunci cand are loc un schimb reciproc de segmente cromozomiale se realizeaza o translocatie reciproca ce necesita doua puncte de ruptura, cate unul la nivelul fiecarui cromozom. Schimbul dintre doi cromozomi neomologi poate fi egal sau inegal, iar detectarea segmentelor implicate se face prin tehnici de bandare. De cele mai multe ori, complementul cromozomial al individului ce contine o translocatie este echilibrat, schimbandu-se numai localizarea genelor pe cromozom. Cand un cromozom pierde prin deletie un fragment ce se va atasa la un alt
|
|
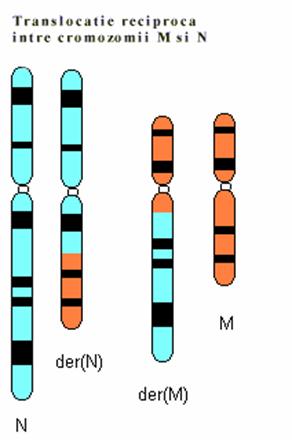
cromozom neomolog fara ca la
cromozomul deletat sa se transfere in schimb un alt fragment se produce o
translocatie nereciproca. In general translocatiile reciproce
intalnite la membrii unei familii sunt caracteristice indivizilor respectivi
desi translocatiile reciproce echilibrate care implica
bratele lungi ale cromozomilor 11 si 22 sunt relativ frecvent
intalnite. Se apreciaza ca incidenta translocatiilor
reciproce este de circa 1/500.
Translocatiile nereciproce au o importanta deosebita in patologia umana deoarece purtatorii fenotipic normali ai unei translocatii echilibrate pot da nastere unor descendenti cu o constitutie cromozomiala neechilibrata. La indivizii cu translocatii reciproce echilibrate, cromozomii nu se pot inperechea in mod normal pentru a forma bivalenti in meioza. Ca urmare, fiecare cromozom se va conjuga cu omologul sau formand un cvadrivalent. Cromozomii din cvadrivalent se pot separa in moduri diferite (fig pag 46).
Sa presupunem ca un individ are o translocatie echilibrata intre bratul lung al cromozomului 8 (8q-) si bratul lung al unui cromozom 12 (12q+). Sansa teoretica a acestui individ de a avea un copil normal este de ¼, ¾ dintre descendenti putand fi purtatori ai unei translocatii echilibrate ca si parintele sau afectati de monosomii sau trisomii partiale.
Translocatia robertsoniana rezulta ca urmare a clivarii a doi cromozomi acrocentrici (13, 14, 15, 21 sau 22) la nivelul centromerului sau in apropierea sa, urmata de fuziunea bratelor lungi. Bratele scurte ale fiecarui cromozom se pierd, acest lucru nefiind de importanta majora pentru organism deoarece ele contin doar gene pentru ARN ribozomal, gene pentru care se gasesc alte numeroase copii la nivelul celorlati cromozomi acrocentrici. Urmarea unei translocatii robertsoniene este reducerea numarului de cromozomi la 45. Deoarece nu se pierde si nici nu se castiga material genetic de importanta majora, translocatia robertsoniana este considerata o restructurare balansata. Incidenta translocatiilor robertsoniene in populatie este
estimata la 1/1000, cea mai frecventa fuziune centrica fiind cea intre bratele lungi ale cromozomilor 13 si 14.
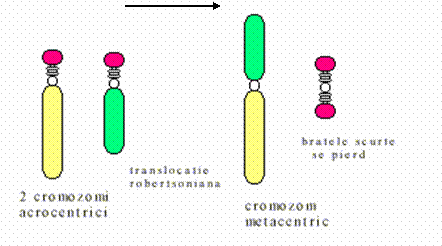
Fig. translocatie robertsoniana
Ca si in cazul translocatiilor reciproce, indivizii cu fuziune centrica pot da nastere unor gameti neechilibrati genetic. De exemplu un purtator a unei translocatii 14q21q va putea produce gameti cu (fig-pag48):
1. Un complement cromozomial normal (cromozomi 14 si 21 normali);
2. Un complement cromozomial balansat (un cromozom 14q21q);
3. Un complement cromozomial neechilibrat ce va contine un cromozom 21 normal si un cromozom 14q21q. Daca acest gamet va participa la fecundare va rezulta un copil cu sindrom Down;
4. Un complement cromozomial neechilibrat caruia ii lipseste cromozomul 21;
5. Un complement cromozomial neechilibrat cu cromozomul 14 lipsa;
6. Un complement cromozomial neechilibrat ce contine un cromozom 14 normal si un cromozom 14q21q.
Ultimele trei tipuri de gameti vor produce, daca vor participa la fecundare zigoti cu monosomie 21, monosomie 14 si trisomie 14. Toate aceste combinatii sunt incompatibile cu viata.
Studiile arata ca femeile cu o translocatie robertsoniena de tipul 13q21q sau 14q21q prezinta un risc de aproximativ 10% de a avea un copil cu sindrom Down in timp ce la barbati riscul este de 1-3%. In cazul purtatorilor unei translocatii robertsoniene 21q21q toti gametii vor fi neechilibrati genetic, fie nulisomici fie disomici pentru cromozomul 21. Ca urmare toti copii ce se vor naste dintr-un cuplu in care unul din indivizi prezinta o astfel de translocatie robertsoniana vor prezenta sindrom Down.
MOZAICISM SI CHIMAERISM
Mozaicismul poate fi definit ca prezenta intr-unul sau mai multe tesuturi ale unui individ a doua sau mai multe linii celulare care difera prin constitutie genetica dar deriva dintr-un singur zigot. Mozaicismul cromozomial rezulta de obicei ca urmare a non-disjunctiei mitotice in primele stadii ale dezvoltarii embrionare urmata de supravietuirea a cel putin doua linii celulare. Daca la a doua diviziune a zigotului cromatidele cromozomului 21 nu se separa vor rezulta patru celule din care doua vor avea 46 de cromozomi, una va contine 47 de cromozomi (trisomie 21) si ultima va contine 45 de cromozomi (monosomie 21). Linia celulara cu 45 de cromozomi are sanse mici de supravietuire si ca urmare embrionul va fi alcatuit din linia celulara normala (circa 66%) si linia trisomica circa 33%. Aproximativ 2-3% din cazurile de sindrom Down sunt datorate mozaicismului.
Mozaicismul poate de asemenea sa existe la nivel molecular daca o mutatie apare de novo intr-o celula somatica sau intr-o linie germinala. Un astfel de tip de mozaicism la nivelul la nivelul unei linii germinale trebuie avut in vedere in acordarea sfatului genetic parintilor sanatosi cu ascendenti sanatosi care au dat nastere unui copil cu distrofie musculara Duchenne.
Himerismul este definit ca prezenta la nivelul unui individ a doua sau mai multe linii celulare diferite din punct de vedere genetic provenite de la mai mult de un zigot. Termenul de himera deriva de la un monstru din mitologia greaca, monstru care avea cap de leu, corp de capra si coada de balaur. La om, se intalnesc doua tipuri de himere: dispermice si sangvine.
Himerele dispermice se formeaza atunci cand doi spermatozoizi diferiti din punct de vedere genetic fertilizeaza doua ovule, rezultand doi zigoti ce vor fuziona dand nastere unui singur embrion. Daca cei doi zigoti au sexe diferite, embrionul himeric va da nastere unui individ cu hermafroditism (cariotip 46, XX/46, XY). Astazi se pot produce in laborator astfel de himere la soarece pentru a facilita studiile transferului de gene.
Himerele sangvine rezulta ca urmare a unui schimb la nivelul placentei intre gemenii dizigoti. De exemplu 90% din celulele unuia din gemeni vor avea cariotipul 46,XY iar eritrocitele vor prezenta predominant grupul de sange B, in timp ce fratele geaman va avea circa 90% din celule cu cariotipul 46, XX si eritrocite ce vor prezenta predominant grupul sangvin A.
NOMENCLATURA SI SIMBOLURI UTILIZATE IN CITOGENETICA
A-G- grupele de cromozomi din cariotip;
1-22- numarul perechilor de autosomi;
46, XX- cariotip normal la femeie;
46, XY- cariotip normal la barbati;
- indica mozaicism, de exemplu 46, XX/47, XXX;
sau : plasarea semnelor inaintea cromozomului indica aditia sau pierderea cromozomului respectiv semnificand astfel o trisomie sau o monosomie; de exemplu 47, XX+21, 45, XX-21. Plasarea semnelor dupa numarul cromozomului indica o trisomie partiala sau o monosomie partiala;
cen centromer;
del deletie;
dup duplicatie;
fra sit fragil;
i izocromozom;
ins- insertie;
inv inversie;
inv.ins. insertie inversa;
ish hibridizare in situ;
p bratul scurt al unui cromozom;
q bratul lung al unui cromozom;
r cromozom inelar;
rep - transloca translocatie reciproca;
rob translocatie robertsoniana;
s satelit;
t translocatie;
ten translocatie in tandem;
ter terminal;
reper cromozomial o trasatura morfologica distincta si constanta care constituie un criteriu important in identificarea cromozomului; reperele include extremitatile bratelor cromozomiale, centromerul precum si o anumita banda;
banda cromozomiala o parte dintr-un cromozom care se distinge clar de partile adiacente printr-o anumita lungime, intunecata sau fluorescenta, colorata sau necolorata in functie de tehnicile folosite;
regiune cromozomiala portiunea dintr-un cromozom cuprinsa intre doua repere adiacente;
In fiecare cromozom regiunile si benzile sunt numerotate de la centromer spre telomere. Cand se desemneaza o banda se indica in ordine numarul cromozomului, simbolul bratului, numarul regiunii si numarul benzii, fara spatii separatoare sau semne de punctuatie. 1p32 indica cromozomul 1, bratul scurt, regiunea 3, banda 2. Prin studiul cromozomilor prometafazici au fost identificate subbenzi cromozomiale care sunt numerotate tot de la centromer spre telomere. Acestea se desemneaza prin plasarea unui punct dupa numarul benzii urmat de numarul subbenzii, de ex. 1p32.3.
Acest sistem de nomenclatura a fost extins pentru a include si rezultatele studiilor FISH. De ex. cariotipul simbolizat 46, XY.ish del(15 q11.2)(D15S10) se refera la un barbat cu o microdeletie la nivelul bratului scurt al cromozomului 15, regiunea 1, banda 1, subbanda 2, identificata prin hibridizare in situ folosind ca proba ADN din locusul D15S10 (D15S10 = ADN din cromozomul 15 situl 10).
ABERATIILE CROMOZOMIALE IN PATOLOGIA UMANA

Pe baza studiilor efectuate se apreciaza ca cca. 10% dintre spermatozoizi si 20% dintre ovocitele mature prezinta un tip sau altul de aberatie cromozomiala. Un procent de 15-20% din totalul sarcinilor sfarsesc printr-un avort spontan iar alti numerosi produsi de conceptie nu supravietuiesc mai mult de cateva zile sau cateva saptamani. Aproximativ 50% din totalul embrionilor avorati spontan) prezinta o aberatie cromozomiala ( tabelul nr..) La nastere acest procent scade la 0,5-1% desi totalul ar fi mai mare daca s-ar lua in considerare si nou-nascutii morti. Aceste observatii indica faptul ca aberatiile cromozomiale sunt raspunzatoare de pierderea unui mare procent din produsii de conceptie umani.
ABERATII NUMERICE ALE AUTOSOMILOR
SINDROMUL DOWN (TRISOMIA 21)
Sindromul Down fost descris de medicul englez J.L. Down inca din 1866 si denumit idiotia mongoloida. Termenul i-a fost sugerat de o superficiala asemanare intre copiii cu acest sindrom si populatiile asiatice. Bazele cromozomiale ale acestui sindrom au fost stabilite insa abia in anul 1959 de catre Lejeune si colaboratorii sai. Datorita faptului ca acest sindrom apare la toate populatiile lumii, iar copii afectati seamana unul cu altul destul de mult, indiferent de populatia careia ii apartin, ca si datorita conotatiilor rasiale, numele de mongoloism a fost inlocuit cu cel de sindrom Down sau trisomia 21.
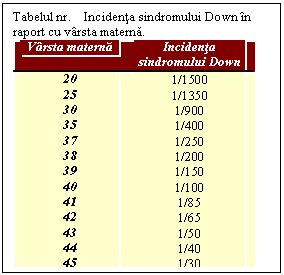 Acest sindrom este datorat in 95% din cazuri
prezentei unui cromozom suplimentar din perechea 21, indivizii
afectati avand cariotipul 47, XX (XY) + 21. (Fig.)Restul
de 5% din indivizi prezinta fie o translocatie fie mozaicism.
Acest sindrom este datorat in 95% din cazuri
prezentei unui cromozom suplimentar din perechea 21, indivizii
afectati avand cariotipul 47, XX (XY) + 21. (Fig.)Restul
de 5% din indivizi prezinta fie o translocatie fie mozaicism.
Dintre toate trisomiile sindromul Down prezinta cea mai larga raspandire, avand o incidenta de 1/650 1/1000 de nou-nascuti din diferite zone geografice (Hook, 1982). Se apreciaza ca din produsii de conceptie cu trisomie 21, 80 sunt eliminati spontan.
Trisomia 21 predomina la baieti (4 baieti: 3 fete). Raportul intre numarul baietilor si al fetelor pare sa nu se modifice chiar dupa introducerea pe scara larga a contraceptivelor orale.
Aparitia acestui sindrom este semnificativ asociata cu varsta inaintata a mamei tabelul nr. ) si cu expunerea la radiatii.
Datorita acestor observatii se apreciaza ca femeile de peste 35 ani reprezinta o grupa cu risc crescut, fapt ce impune indicatia de analiza citogenetica prin amniocenteza.
Fig. nr. Cariotip 47XXY+21
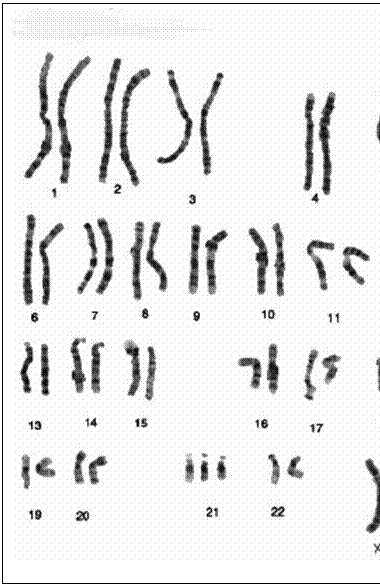
Diagnostic
clinic
Nici unul din semnele clinice ale sindromului nu este patognomic, dar ansamblul lor este caracteristic. In majoritatea cazurilor, diagnosticul clinic se poate pune de la nastere: copilul este hipoton, deseori prezinta hiperlaxitate ligamentara, pielea este catifelata si marmorata.
Simptomele caracteristice ale sindromului Down la adult si la nou nascut sunt prezentate in tabelul de mai jos.
 Aceste semne majore se accentueaza sau
diminueaza cu varsta. Exista numeroase variatii individuale.
Aceste semne majore se accentueaza sau
diminueaza cu varsta. Exista numeroase variatii individuale.
Dismorfia cranio-faciala
Craniul este mic si rotund. Diametrul anteroposterior este redus, iar occipitul este aplatizat. Indicele cefalic, ale carui valori normale sunt de 0,75 -0,80, este mai mare decat 0,83. Fontanelele sunt mari si se inchid cu intarziere. Fata este rotunda, profilul facial este plat datorita hipoplaziei oaselor proprii ale nasului, iar fruntea este bombata.
Fantele palpebrale sunt orientate oblic in sus si in afara. Aproximativ 50% din pacienti prezinta pliuri epicantice, care mascheaza unghiul intern al ochiului. Strabismul convergent, blefaritele si nistagmusul sunt simptome relativ frecvente. Din cauza unor defecte de dezvoltare, irisul are un aspect patat (petele Brushfield) in 75% din cazuri.
Aceste pete mici, albicioase, rotunde sau neregulate formeaza o coroana situata la jonctiunea treimii mijlocii cu treimea externa a irisului. Opacifierea cristalinului este observata la peste 50% din pacienti. Fantele palpebrale sunt inguste, ingustimea lor fiind legata de hipogeneza orbitelor si de volumul redus al globilor oculari. Infectiile conjunctivei palpebrale sunt frecvente si persistente si duc de multe ori la disparitia genelor.
Urechile sunt mici si rotunde. Helixul are un aspect rulat, iar antehelixul este proeminent. Lobul urechii este mic si aderent. Conductul auditiv este mic, iar hipoacuzia sau surditatea reprezinta simptome comune.
Gura este mica si aproape permanent intredeschisa. Buza inferioara este groasa, eversata, aproape permanent umeda. Maxilarul superior este ingust si foarte scurt. Cavitatea bucala este mica datorita hipogenezei etajului inferior al scheletului facial.
Proeminenta limbii in afara cavitatii bucale este unul din semnele majore ale sindromului. Pana de curand s-a considerat ca trisomicii 21 prezinta o macroglosie adevarata.) S-a constatat insa ca este vorba de o pseudomacroglosie (limba proeminand in afara datorita cavitatii bucale mici si a mandibulei hipoplazice). Limba are deseori aspect scrotal tipic, uneori prezinta leziuni exfoliative.
Volumul mare al amigdalelor, ca si displazia foselor nazale, explica disfunctia respiratorie cronica a pacientilor cu sindrom Down. Bolta palatina este, de obicei, ogivala. Se remarca uneori absenta luetei. Dintii sunt fragili, erup tarziu, sunt vicios implantati. Anizodontia este obisnuita. Cu toate acestea cele doua dentitii pot aparea.
Toate aceste anomalii: hipoplazia cavitatii bucale, endognatia maxilarului superior si implantarea vicioasa a dintilor, explica deficientele de masticatie. Trisomicii 21 sug si mesteca defectuos, uneori nu pot ingera decat alimente semilichide sau tocate.
Gatul este scurt si gros.
|
Politica de confidentialitate |
| Copyright ©
2026 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |